סטראמנופילס
ה-סְטְרַמֵנוֹפִילס (Stramenopiles לעיתים נקראת באנגלית Heterokont) היא הקבוצה הגדולה ביותר בתוך על-קבוצת SAR וקבוצה משמעותית בתוך האיקריוטים. מקובל להגיד שהיא מכילה כ-25,000 מינים ידועים.[1] רוב המינים של קבוצה זאת נחשבים לאצות והם מכילים מגוון רב של צורות חיים ומבנים מורפולוגיים, מיצורים רב-תאיים ענקיים כמו הקלפ (ממחלקת החומיות) ועד יצורים דו-תאיים או חד-תאיים כמו הצורניות. חלק מהמגוון של הקבוצה מתבטא גם בעובדה שחלק מהמינים אוטוטרופים (כמו החומיות והצורניות) וחלק מהמינים הטרוטרופים (כגון הסדרה Oomycete המכילה מינים טפיליים רבים כמו ה-Phytophthora שגרמה לרעב תפוחי האדמה הגדול באירלנד). למרות המגוון הרב, מקובל לראות בסטרמנופילים קבוצה מונופילטית.[2]

| |
| מיון מדעי | |
| על־ממלכה: | איקריוטים |
| קבוצה: | על-קבוצת SAR |
| קבוצה: | סטראמנופילס |
| שם מדעי | |
| שמות נוספים | |
|
Heterokonta | |

על פי ג. ק. דוד[3] מקור השם הוא בשם העצם היווני Straw, שמשמעותו קש, הרומז על השערות היוצאות מהשוטונים של חלק מהמינים. מקור השם Heterokont, נגזר מהמילה היוונית έτερος (תעתיק: הטרוס) שמשמעותה היא "שונה", הוא בחלוקה על פי תומאס קלבייר-סמית.[4] שם זה, הנהוג גם לפעמים בספרות, נגזר מהעובדה שלמינים בעלי השוטונים ישנם שוטונים בעלי אורך שונה.
מורפולוגיה
עריכה
מכיוון שמדובר בקבוצה ששורשיה האבולוציונים הם קדומים ביותר, יש מעט מאפיינים פיזיולוגים ברורים שניתן לשייך לכל המינים בתוכה, מכיוון שרבים מהמאפיינים האלה הופיעו או נעלמו במהלך הברירה הטבעית. למרות זאת, ישנן תכונות הקיימות בלפחות חלק מהמינים בכל אחת מהקבוצות ולכן הן נחשבות לתכונות מאחדות של כלל המינים.
תאים בעלי כושר תנועה
עריכהרבים מהסטרמנופילים הם חד-תאיים בעלי שוטונים. חלק מהמינים חסרי שוטונים ברוב שלבי החיים שלהם אך יפתחו שוטונים בשלב חיים מסוים, בדרך כלל בשלב הגמטות. לרוב, יהיו לתאים אלה שני שוטונים, אשר יהיו נבדלים באורכם בצורה ברורה, ממאפיין פיזיולוגי ברור זה נגזר גם השם הנרדף Heterokont. השוטון הארוך של תאים אלה מכוסה בשורה או שתיים של שערות הנקראות מסטיגונמות (באנגלית: mastigonemes) המחולקות בצורה ברורה לשלושה חלקים,[5] ולעיתים נדירות לשני חלקים. השוטון הקצר של תאים אלה הוא בדרך כלל חלק ולעיתים כמעט ולא בולט מתוך הגוף המרכזי. לעיתים יהיה מבנה של מיקרו-צינוריות התומך בשוטונים בבסיסם.
המסטיגונמות מיוצרות משרשראות של גליקופרוטאין בתוך הרשתית התוך-פלזמית[6] לפני שהן מוסעות בתוך גוף התא אל מקומם על החלק החיצוני של השוטון. כאשר השוטון זז, השערות יוצרות זרם הופכי לכיוון התנועה ומושכות את התא בתוך המים[6] ובמקרים מסוימים גם עוזרות לו לאסוף אוכל.[7] העובדה שהשערה מחולקת באופן ברור לשלושה חלקים אופיינית לסטרמנופילים ונמצאת במספר מועט של קבוצות מחוץ לקבוצה זאת (בצורה בולטת בעיקר ב-Cryptista[8] שמעמדה הסיסטמטי לא ברור) אם כי חשוב לציין שכמה מהשושלות בתוך הקבוצה, כמו הצורניות, איבדו את התכונה המאפיינת הזאת.
כלורופלסטים
עריכהלהרבה מהסטרמנופילים יש כלורופלסטים המוקפים בארבע שכבות של ממברנות, דבר המיחד אותם מהכלורופלסטים של שאר הקבוצות האיקריוטיות השונות. הממברנה הפנימית ביותר היא למעשה המשך של הרשתית התוך-פלזמית, הממברנה השנייה מבדילה בין הרשתית לבין הנהור. לאחר מכן יבואו שתי ממברנות צמודות שביניהן ימצא הקרום של התילקואיד. הסידור הזה של ארבע ממברנות מייצג לדעת חוקרים רבים את העובדה שמקור הכלורופלסטים אצל הסטרמנופילים הוא באנדוסימביוזה של אדומית בתוך אחד מהאבות הקדומים של כל הסטרמנופילים שמבצעים פוטוסינתזה.[9][10]
לכלורופלסטים של הסטרמנופילים יש בדרך כלל גם כלורופיל a וגם כלורופיל b ובדרך כלל הם מכילים בעיקר את הפיגמנט פוקוקסנטין[11] המעניק לקבוצות שונות את צבען החום-ירקרק.
מיון
עריכהלמרות המאפיינים הפיזיולוגים המשותפים של רבים מהסטרמנופילים, אפיון מלא של הקבוצה כקבוצה מונופילטית התרחש רק לאחר מחקרים פילוגנטיים.[12] כיום נהוג למקם את הסטרמנופילים בתוך על-קבוצת SAR, שהיא בעיקרה קבוצה שקשה להצביע על מאפיינים פיזיולוגים דומים של כל הנמצאים תחתה.
מיון פנימי
עריכהנהוג לחלק את הסטרמנופילים מורפולוגית לשלוש קבוצות עיקריות:[13]
- Ochrophyta (דמויי האצות) - מערכה זאת מכילה את כל המחלקות שמבצעות פוטוסינתזה ומכילות כלורופיל. מקובל כיום לראות במערכה זאת קבוצה מונופילטית.[14] מכילה לדוגמה את הצהוביות, החומיות והצורניות.
- Pseudofungi (דמויי פטריות, באנגלית נקראים גם לפעמים: Heterokontimycotina) - מכילה קבוצות המזכירות בצורת החיים שלהם פטריות, אך רחוקות אבולוציוניות מהן. מקובל לראות היום בקבוצה זאת קבוצה פוליפילטית. המינים בקבוצה זאת הם כולם הטרוטרופים.
- Bigyra - קבוצה המכילה את כל שאר המחלקות שאינן נמצאות בקבוצות האחרות. קבוצה זאת היא פרפילטית.
נעשו בשנים האחרונות מספר חלוקות[15][16] המנסות למדל בצורה יותר טובה את החלוקה הפילוגנטית בתוך הסטרמנופילים אך חלוקות אלה עדיין שנויות במחלוקת ומשתנות באופן תדיר.
מגוון פנימי
עריכהבגלל עושר המינים הרב בתוך הקבוצה, קיים בתוכה מגוון רב של צורות חיים וצורות פיזיולוגיות.
-
 תפוחי אדמה נגועים ב-Phytophthora.
תפוחי אדמה נגועים ב-Phytophthora. -
 Synura, אצת מים מתוקים בעלת מבנה מיוחד.
Synura, אצת מים מתוקים בעלת מבנה מיוחד. -

-
 צורנית - אצה פלנקטונית המבצעת פוטוסינתזה.
צורנית - אצה פלנקטונית המבצעת פוטוסינתזה. -
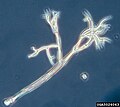 Peronospora - שייכת לדמויי-הפטריה, ניתן לראות את המבנה המזכיר את התפטיר.
Peronospora - שייכת לדמויי-הפטריה, ניתן לראות את המבנה המזכיר את התפטיר. -
 אצה מהזהוביות.
אצה מהזהוביות.






קישורים חיצוניים
עריכה- סטראמנופילס, באתר NCBI (באנגלית)
- סטראמנופילס, באתר האנציקלופדיה של החיים (באנגלית)
הערות שוליים
עריכה- ^ Chapman, A. D., Numbers of Living Species in Australia and the World, 2009 (ארכיון)
- ^ Michael A. Ruggiero, Dennis P. Gordon, Thomas M. Orrell, Nicolas Bailly, A Higher Level Classification of All Living Organisms, PLOS ONE 10, 2015-04-29, עמ' e0119248 doi: 10.1371/journal.pone.0119248
- ^ Constancea 83.16: Names of Fungi above the Rank of Order, ucjeps.berkeley.edu
- ^ Eukaryote kingdoms: Seven or nine?, ResearchGate (באנגלית)
- ^ Laura Barsanti, Paolo Gualtieri, Algae: Anatomy, Biochemistry, and Biotechnology, CRC Press, 2005-11-14. (באנגלית)
- ^ 1 2 G. Benjamin Bouck, The Structure, Origin, Isolation, and Composition of the Tubular Mastigonemes of the Ochromonas Flagellum, The Journal of Cell Biology 50, 1971-08-01, עמ' 362–384 doi: 10.1083/jcb.50.2.362
- ^ THEODORE L. JAHN, MICHAEL D. LANOMAN, JAMES R. FONSECA, The Mechanism of Locomotion of Flagellates. II. Function of the Mastigonemes ofOchromonas*, The Journal of Protozoology 11, 1964-08, עמ' 291–296 doi: 10.1111/j.1550-7408.1964.tb01756.x
- ^ D.E. Bradley, The ultrastructure of the flagella of three chrysomonads with particular reference to the mastigonemes, Experimental Cell Research 41, 1966-01, עמ' 162–173 doi: 10.1016/0014-4827(66)90556-8
- ^ D. L. Robertson, Evolution of Glutamine Synthetase in Heterokonts: Evidence for Endosymbiotic Gene Transfer and the Early Evolution of Photosynthesis, Molecular Biology and Evolution 23, 2006-02-22, עמ' 1048–1055 doi: 10.1093/molbev/msj110
- ^ Jim Provan, Susan Murphy, Christine A Maggs, Universal plastid primers for Chlorophyta and Rhodophyta, European Journal of Phycology 39, 2004-02, עמ' 43–50 doi: 10.1080/09670260310001636668
- ^ Niels Daugbjerg, Peter Henriksen, PIGMENT COMPOSITION AND rbcL SEQUENCE DATA FROM THE SILICOFLAGELLATE DICTYOCHA SPECULUM: A HETEROKONT ALGA WITH PIGMENTS SIMILAR TO SOME HAPTOPH Y TES, Journal of Phycology 37, 2001-12, עמ' 1110–1120 doi: 10.1046/j.1529-8817.2001.01061.x
- ^ J. H. Gunderson, H. Elwood, A. Ingold, K. Kindle, Phylogenetic relationships between chlorophytes, chrysophytes, and oomycetes, Proceedings of the National Academy of Sciences 84, 1987-08-01, עמ' 5823–5827 doi: 10.1073/pnas.84.16.5823
- ^ Ingvild Riisberg, Russell J.S. Orr, Ragnhild Kluge, Kamran Shalchian-Tabrizi, Seven Gene Phylogeny of Heterokonts, Protist 160, 2009-05, עמ' 191–204 doi: 10.1016/j.protis.2008.11.004
- ^ Thomas Cavalier-Smith, Ema E-Y. Chao, Phylogeny and Megasystematics of Phagotrophic Heterokonts (Kingdom Chromista), Journal of Molecular Evolution 62, 2006-03-22, עמ' 388–420 doi: 10.1007/s00239-004-0353-8
- ^ Michael A. Ruggiero, Dennis P. Gordon, Thomas M. Orrell, Nicolas Bailly, A Higher Level Classification of All Living Organisms, PLOS ONE 10, 2015-04-29, עמ' e0119248 doi: 10.1371/journal.pone.0119248
- ^ Thomas Cavalier-Smith, Josephine Margaret Scoble, Phylogeny of Heterokonta: Incisomonas marina, a uniciliate gliding opalozoan related to Solenicola (Nanomonadea), and evidence that Actinophryida evolved from raphidophytes, European Journal of Protistology 49, 2013-08, עמ' 328–353 doi: 10.1016/j.ejop.2012.09.002