מערכת הראייה
מערכת הראייה היא מכלול איברי החישה והמסלולים העצביים המאפשרים לבעלי חיים לראות. מערכת זו קולטת מידע חזותי, אשר מקורו באור המגיע לעין, ומגבשת בעזרתו תפיסה של הסביבה.
_(20383247785).jpg) | |
| שיוך |
תחושה, heterogeneous anatomical cluster, ישות אנטומית מסוימת |
|---|---|
| מיקום אנטומי |
עין |
| מזהים | |
| FMA |
7191 |
| מערכת השפה הרפואית המאוחדת |
C3536733 |
המשימה המוטלת על מערכת הראייה מורכבת למדי – יצירת דימוי של עולם תלת-ממדי על סמך היטלים דו-ממדיים רגעיים שלו על רשתיות העיניים. בבני אדם, מערכת הראייה היא המפותחת במערכות החושים ומבחינות רבות ביצועיה עולים על אלו של מערכות הראייה הממוחשבת המתקדמות ביותר.
מערכת הראייה מסתמכת על תהליך של התמרה חזותית (אנ'), שבו פוטונים שפוגעים ברשתית עוברים התמרה לאותות חשמליים של מערכת העצבים.
מערכת הראייה המתוארת להלן קיימת ביונקים, אם כי רבים ממרכיביה משותפים גם לבעלי גולגולת אחרים.
מערכת הראייה האנושית
עריכה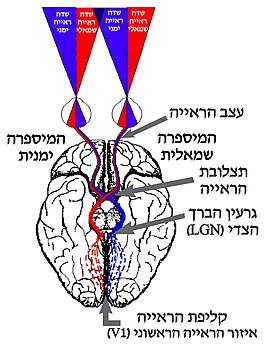
מערכת הראייה משלבת בתוכה את העין, שהיא איבר החישה שקולט את הגירוי הראייתי, ואת כל האזורים והרשתות העצביות במוח שתפקידם הוא לעבד את הגירוי. בנוסף לעיבוד המידע החזותי, המוח שולט באופן פעיל על כניסתם של גירויים למערכת, וזאת באמצעות הכוונה של תנועות העיניים וויסות הקשב החזותי.
ברשתית העין מתבצעת ההתמרה של האור לאותות חשמליים, על מנת שניתן יהיה לייצג את המידע בתוך מערכת העצבים. מתוך הרשתית יוצאים אקסונים של תאי עצב, שלאורכם עובר המידע על הגירוי הראייתי. מידע זה מועבר אל המוח דרך עצב הראייה, ומשם מתפצל למספר מסלולים אשר מערבים גרעינים שונים בתוך המוח. במסלולים אלה מתבצע עיבוד מתקדם של המידע החזותי, אשר ישפיע בסופו של דבר על ההתנהגות.
במסלול העצבי העיקרי של המידע החזותי, האקסונים ממשיכים דרך צרור הראייה (Optic tract), שהוא המשכו של עצב הראייה, אל התלמוס, שבו אזור הנקרא גרעין הברך הצדי. באזור זה מתבצע העיבוד הראשוני של המידע החזותי. בעוברם דרך עצב הראייה, כמחצית מהאקסונים חוצים לצדו הנגדי של הראש במקום שנקרא תצלובת הראייה (Optic chiasm). בצורה זו, המידע שמגיע משתי העיניים מתפצל בחלוקה לפי שני שדות ראייה: ימני ושמאלי. מן גרעין הברך הצדי יוצא צרור סיבים עצביים של תאי העצב שלו, הפרוש לאורך אזור הנקרא מַעֲנִית הַפָּרִישׁ ומוליך את האותות אל אזורים בקליפת המוח הראייתית, אשר נמצאת באונה העורפית. באזורים אלה מתבצע עיבוד מתקדם יותר של המידע החזותי. מסלול זה הוא החשוב ביותר מבחינת עיבוד המידע החזותי, והוא הכרחי ליצירת תפיסה חזותית מודעת.
המסלול העצבי השני עובר מצרור עצבי הראייה אל התליל העליון (Superior colliculus) של הטקטום במוח התיכון. אזור זה מבקר את תנועות שרירי גלגל העין ומחווט אף הוא אל קליפת המוח, דרך הכר התלאמי (Pulvinar). ככל הנראה מועבר מידע חזותי לקליפת המוח גם במסלול משני זה. תנועת שרירי גלגל העין מבוקרת גם באופן בלתי רצוני גם על ידי הרפלקס הפרוזדורי-עיניתי (Vestibulo-ocular reflex), השומר על עקיבת העיניים אחרי עצמים בזמן תנועות הראש. רפלקס מהיר זה, החיוני למשל לקריאה, מתבסס על מידע המגיע מהאוזן הפנימית בנוגע לשיווי המשקל הרגעי של הראש.
המסלול העצבי השלישי עובר מתאי גנגליון קולטי-אור ברשתית (Photosensitive ganglion cell), דרך הצרור הרשתיתי-היפותלמי (Retinohypothalamic tract) אל הגרעין העל-תצלובתי (Suprachiasmatic nucleus) בהיפותלמוס. מסלול זה משמש לתיאום המקצב היממתי של השעון הביולוגי ביחס לרמות התאורה השוררות בסביבה, על ידי תזמון ייצורם והפרשתם של הורמונים בהיפותלמוס ובבלוטת יותרת המוח, שמווסתים תפקודי ערות שונים. כמו כן הם שולחים את סיביהם לעבר הפרטקטום (Pretectum) שבמוחה הקדמית (Prosencephalon), השולט על שרירי הקשתית המווסתים את קוטר האישון[1]. מקצת הסיבים העצביים מתאי גנגליון קולטי-אור אלה נשלחים לעבר התליל העליון ואף לעבר גרעין הברך הצדי[2][3], אך כיום עדיין לא ידוע אם הם מעורבים ישירות בעיבוד התפיסה החזותית של העולם.
העין
עריכה
 ערך מורחב – עין
ערך מורחב – עין
העין היא איבר מורכב למדי. קיים דמיון חיצוני מסוים בין העין האנושית לבין המצלמה, שמחקה את פעולת העין באופן חלקי. גם בעין וגם במצלמה נעשה שימוש בעדשה שקופה למיקוד האור לעבר מערך של קולטנים רגישים לאור ברשתית העין, או התקן רגיש לאור כמו סרט צילום במצלמה אנלוגית או חיישן אלקטרוני במצלמה דיגיטלית. גם צמצם המצלמה דומה לקשתית העין בהיותו התקן טבעתי המווסת את שטף האור המגיע לרשתית. עם זאת, אופן הפעולה של העין שונה מהרבה בחינות אחרות מזה של כל מצלמה, בהתאם לתפקידים השונים שהיא ממלאת.
האור המגיע לעין נשבר על ידי הקרנית. בהמשך הוא עובר דרך הנקב במרכז קשתית העין, המכונה אישון. עדשת העין ממקדת את קרני האור, לקבלת דמות ממשית הפוכה על הרשתית.
הרשתית
עריכה- ערך מורחב – רשתית

האור עובר דרך שכבות אלו (משמאל לימין) עד שהוא פוגע בתאים קולטי אור (השכבה הימנית ביותר). בעקבות תגובה פוטוכימית בתוך התאים קולטי האור, מתקדם אות חשמלי לתאים הדו-קוטביים (מסומנים באדום) והאופקיים (בשכבה הצהובה). משם מתקדם האות לתאי האמקרין והגנגליון. המידע מועבר אל המוח דרך האקסונים של תאי הגנגליון, שמרכיבים את עצב הראייה.
הרשתית היא החלק בעין שקולט את האור ומתמיר אותו לדחפים עצביים. הרשתית בנויה כמערך רבוד של שכבות תאים, כאשר תהליך ההתמרה(אנ') מתבצע בתאים קולטי אור אשר נמצאים בשכבה האחורית. האור, אשר מוקד על ידי עדשת העין, חולף דרך שתי שכבות של תאי עצב בתוך הרשתית לפני שהוא פוגע בתאים קולטי האור. לאחר התהליך שמתרחש בתוכם, האותות המותמרים מועברים חזרה דרך אותם תאי עצב, אשר מבצעים עיבוד ראשוני של המידע ומעבירים אותו הלאה אל המוח. מסלול ה-phototransduction(אנ')[4] הוא כלל התהליכים המעורבים בראייה מרמת התאים הרגישים לאור, ועד יציאת סיגנל חשמלי לעצב הראייה. לתהליך העיבוד שמתרחש בתוך הרשתית ישנה חשיבות לאופי התפיסה הראייתית – אינטראקציות בתוך קבוצות של תאי עצב ברשתית מובילות לכך שהאות שמועבר החוצה אינו סכום ליניארי פשוט של המידע הראייתי שנקלט.
התמרת האור לאותות עצביים
עריכה- ערך מורחב – תא קולט אור

.
התאים קולטי האור ברשתית ממירים את האור שנכנס לעין לאותות חשמליים. תאים אלה מכילים חלבונים ממברנליים ממשפחת האופסינים, שהם קולטנים לאור (פוטורצפטורים).
כל האופסינים מכילים קבוצה פונקציונלית בשם "רטינל", המשנה את תצורתה המרחבית כאשר היא בולעת פוטון יחיד של אור. שינוי התצורה מוביל לשרשרת ביוכימית של מעבר אותות. בסוף התהליך, התא קולט האור עובר היפרפולריזציה, ובכך משנה את האותות החשמליים המועברים הלאה לתאי העצב שאליהם הוא מחובר. תאים קולטי אור שומרים על מתח מנוחה גבוה יחסית, שדי בו כדי לגרום לשחרור מתמיד של מוליך עצבי. תהליך ההיפרפולריזציה שמתרחש בהשפעת האור מעכב את שחרור המוליך העצבי אל הסינפסות עם תאי העצב.
מידת הרגישות של הקולטן לתדרים אלקטרומגנטיים שונים (כלומר, לפוטונים באורכי גל שונים) נקבעת על ידי החלבון הייחודי הקשור לרטינל. נכון לשנת 2005, זוהו למעלה מאלף חלבונים שונים במשפחת האופסינים[5], כאשר שישה מתוכם קיימים ברשתית האנושית. בהתאם לתפקודם ולסוג האופסינים שהם מכילים, התאים קולטי האור ברשתית מתחלקים לשלוש קבוצות:
- קנים (Rods) – מכילים את החלבון רודופסין. תאים אלה הם רגישים מאוד לאור, ויכולים להיות מופעלים על ידי פוטונים בודדים. בעזרתם מתאפשרת ראייה גם בתנאים של הארה חלשה ביותר.
- מדוכים (Cones) – מכילים פוטופסינים. בבני אדם קיימים לרוב שלושה סוגים של מדוכים, אשר כל אחד מכיל וריאציה שונה של הקולטן. תאים אלה משמשים לראייה בסביבה מוארת, והם מאפשרים להבחין בין צבעים שונים.
- תאי גנגליון קולטי אור – מכילים את הקולטן מלאנופסין. הפלט מתאים אלה מסייע להתאים את קוטר האישון והמקצב היממתי של השעון הביולוגי לרמות ההארה השוררות בסביבה[6][1][2][3].
בסך הכול יש בעין כשישה מיליון מדוכים וכ-125 מיליון קנים. אלה מצופפים על גזרה של 72% משפת כדור שקוטרו 22 מ"מ, באדם בוגר. במרכז הרשתית נמצאת נקודת היציאה של האקסונים אל עצב הראייה, אשר מכונה "הכתם העיוור" (Blind spot) היות שאין בה תאים קולטי אור. זהו אזור אליפטי שצבעו לבן, בשטח כ-3 מ"מ רבועים. מצד הכתם העיוור, בכיוון הרקות נמצא הכתם שבמרכזו הגומה המרכזית (Fovea). הגומה המרכזית מרוצפת בכוורת צפופה במיוחד של מדוכים קטנים ביותר, בסידור משושה המאפשר ניצולת מרבית של פני השטח ללא שטחים מתים. הצפיפות המשטחית של המדוכים בגומה המרכזית מגיעה לכ-140,000 מדוכים למילימטר רבוע. כך מושגת חדות ראייה מרבית בגומה המרכזית, על חשבון רגישות פחותה לאור קלוש.
ככל שמתרחקים מן הגומה המרכזית לכיוון שולי הרשתית, צפיפות המדוכים יורדת ושכיחות הקנים עולה. בזווית של כעשרים מעלות ביחס לקו הראייה מגיעה צפיפות הקנים לשיא של כ-160,000 קנים למילימטר רבוע. היות ששיעור קטן יותר של תאי גנגליון מוקדשים לעיבוד המידע משולי הרשתית, רגישות העין לאור היא מרבית בהיסט קטן יותר: 8–16 מעלות ביחס לקו הראייה. אסטרונומים המחפשים גרמי שמיים חיוורים משתדלים להביט מעט הצידה ממיקומם המשוער, כדי להיעזר ברגישות המוגברת של העין לאור בזוויות אלה. שיטה זו נקראת ראייה מופנית.
תאי עצב בתוך הרשתית
עריכהבתוך הרשתית קיימות מספר אוכלוסיות של תאי עצב, אשר בהן מתבצע העיבוד הראשוני של המידע הראייתי. מהתאים קולטי האור, האותות העצביים עוברים לתאי רשתית דו-קוטביים, ומשם לתאי גנגליון, שהם אלה שמעבירים את המידע החוצה דרך עצב הראייה. בנוסף, הרשת העצבית הזו משלבת בתוכה שני סוגים נוספים של תאי עצב, הנקראים תאים אופקיים ותאי אמקרין.

נסתכל על תא גנגליון מסוג 'מרכז-דולק' ששדה הקלט שלו סמוך לגבול בין שני פסים. כאשר שדה הקלט הוא בקצה השמאלי של הפס, חלק מהיקף שדה הקלט נופל על אזור כהה יותר, ולכן התא יגיב בצורה יחסית חזקה וייצור תפיסה של אזור שהוא יותר בהיר. באופן דומה, כאשר שדה הקלט נמצא בקצה הימני של הפס, חלק מהיקף שדה הקלט נופל על אזור מואר יותר, ודבר זה מקטין את תגובת התא ויוצר תפיסה של נקודה כהה יחסית.
מאפייני העיבוד הראייתי ברשתית
עריכהבהתאם לאוכלוסיות של התאים קולטי האור שמהם הם מקבלים קלט עצבי, לתאי העצב ברשתית ישנם שדות קלט מוגדרים. משמעות הדבר היא שדפוס הדחפים העצביים של כל תא עצב מושפע אך ורק מגירויים ראייתיים שנמצאים באזור מסוים של שדה הראייה, שהוא שדה הקלט של אותו תא. כך, בסיכום של כלל התאים ברשתית, נוצר מיפוי עצבי של שדה הראייה השלם.
פעולתה של הרשת העצבית ברשתית יוצרת במרבית תאי הגנגליון דגמי הפעלה אופייניים, שבהם התגובה לאור היא שונה בחלק המרכזי של שדה הקלט של התא לעומת ההיקף של שדה הקלט. בחלק מהתאים, תא הגנגליון מגיב בצורה מרבית למצב שבו קיימת הארה במרכז שדה הקלט שלו אך לא קיימת הארה בהיקף של שדה הקלט. תאים אלה מכונים תאי "מרכז-דולק" (ON-center). באופן דומה, בתאים מסוג "מרכז-כבוי" (OFF-center), התא מגיב בצורה מרבית כאשר ישנה הארה בהיקף שדה הקלט, אך לא קיימת הארה במרכזו. בתאי גנגליון אחרים, ההבחנה בין המרכז וההיקף של שדה הקלט מבוססת לא על רמת הארה, אלא על צבעים שונים – למשל, קיימים תאים שמגיבים בצורה חזקה לאור אדום במרכז שדה הקלט שלהם לצד אור ירוק בהיקף שדה הקלט.
דפוס ההפעלה של תאי הגנגליון בא לידי ביטוי באופן שבו אנו תופסים גירויים חזותיים. ההבחנה בין מרכז להיקף בשדות הקלט יוצרת רגישות גבוהה לניגודיות, שכן העיבוד העצבי שם דגש על מצבים שבהם יש הבדל בהארה או בצבע בין נקודה מסוימת לבין הסביבה הקרובה שלה, ולאו דווקא על עוצמת ההארה המוחלטת.
מבנה הרשת העצבית
עריכההמדוכים והקנים יוצרים סינפסות עם תאים דו-קוטביים, שמעצבבים בתורם את תאי הגנגליון.
ברשת תאי העצב ברשתית מתבצעת דחיסה של הקלט הראייתי: בעוד ישנם כ-130 מיליון תאים קולטי אור, עצב הראייה מכיל רק כ-1.2 מיליון אקסונים של תאי גנגליון. עם זאת, רמת הדחיסה והסינון של המידע משתנה מאוד בהתאם למיקום על פני הרשתית. בגומה המרכזית, שבה מושגת חדות הראייה המרבית, אין כלל דחיסה של הקלט – על פי הערכות, על כל תא מדוך בגומה המרכזית באדם ישנם 2 תאי גנגליון לפחות[7].
בדומה לתאי הגנגליון, גם התאים הדו-קוטביים שמעצבבים אותם נחלקים לתאים מסוג 'דולק' (ON) ו'כבוי' (OFF). חלוקה זו מתבססת על תגובתם של התאים למוליך העצבי שמשחררים התאים קולטי האור. כאמור, כאשר אור פוגע בתא קולט אור, התא עובר היפרפולריזציה. דבר זה מוריד את כמות הגלוטמט שמופרש לתאים הדו-קוטביים. בתאים דו-קוטביים מסוג 'דולק', הירידה בכמות הגלוטמט תוביל לדפולריזציה, ואז התא יעביר אות מעורר לתאי הגנגליון. בתאים מסוג 'כבוי', הירידה בשחרור הגלוטמט תיצור היפרפולריזציה, ותעכב את העברת האותות הלאה.
המבנה המיוחד של שדות הקלט בתאי הגנגליון, שבו יש הפרדה בין מרכז להיקף, מתבססת על פעולתם של התאים האופקיים, אשר יוצרים מצב של "עיכוב צידי" (Lateral Inhibition). התאים האופקיים מקבלים קלט מהתאים קולטי האור, אך במקביל הם גם מווסתים את הפעילות של תאים אלה. אופן הסידור של תאי העצב הוא כזה שבו התאים הדו-קוטביים מקבלים קלט אך ורק מתא קולט אור שנמצא במרכז שדה הקלט שלהם, ואילו התאים האופקיים מקושרים לכל התאים קולטי האור באותו שדה קלט. במצב זה, התאים האופקיים מסוגלים לקבל מידע על התאורה בהיקף שדה הקלט, ולהשתמש במידע זה על מנת להשפיע על פעילותו של התא קולט האור שבמרכז שדה הקלט ובכך על המידע שמועבר הלאה לתאים הדו-קוטביים ולתאי הגנגליון.
מערכת זו פועלת באופן הבא: התאים האופקיים מפרישים באופן קבוע את המוליך העצבי GABA לתאים קולטי האור. ירידה בכמות הגלוטמט שמופרש מהתא קולט האור, שמתרחשת בעקבות פגיעת אור בקולטן, גורמת לתא האופקי לעבור היפרפולריזציה ולשחרר פחות GABA. הירידה בשחרור ה-GABA גורמת בתורה לדפולריזציה של התאים קולטי האור שמקבלים קלט מהתא האופקי, ובכך מעודדת העברת סיגנל של 'חושך'. עם זאת, היחס המספרי בין התאים קולטי האור בהיקף שדה הקלט לבין התא הבודד שבמרכזו גורם לכך שהתא האופקי ישפיע על העברת האותות מהתא המרכזי רק אם קיימים אותות שונים מהתאים ההיקפיים. אם, למשל, רק התא שבמרכז קלט אור ועבר היפרפולריזציה, ההשפעה על התא האופקי תהיה קטנה יחסית והאפקט המווסת שלו לא יספיק על מנת לגרום לדפולריזציה בתא קולט האור. לעומת זאת, אם ייקלט אור במספר תאים שנמצאים בהיקף שדה הקלט, תהיה ירידה יחסית גדולה בשחרור ה-GABA מהתא האופקי, והיא תספיק על מנת לגרום לדפולריזציה משמעותית בתא קולט האור במרכז שדה הקלט.
תאי אמקרין נמצאים בשכבה של הקישור בין התאים הדו-קוטביים לתאי הגנגליון. אף על פי שתאים אלה אחראים לכ-70% מהקלט שמגיע לתאי הגנגליון, ידוע מעט יחסית על תפקידם. במקביל להשפעתם על תאי הגנגליון, תאי האמקרין מווסתים את התאים הדו-קוטביים על סוגיהם השונים. ישנם עשרות סוגים של תאי אמקרין, אשר מתמחים ככל הנראה בהיבטים שונים של ראייה ורגישות לאור. הסוגים השונים של תאי האמקרין נבדלים במבנה, בהיקף הקישוריות שלהם ובאופן פעולתם, כאשר לרובם ישנה השפעה מעכבת דרך המוליכים העצביים GABA או גליצין.
עיבוד הקלט הראייתי בתוך המוח
עריכה
מעבר דרך התלמוס
עריכהכאמור, במסלול העיקרי לעיבוד הקלט הראייתי המידע עובר מתאי הגנגליון ברשתית אל גרעין הברך הצדי (LGN) בתלמוס. גרעין הברך הצדי בהמיספרה הימנית של המוח מקבל מידע מהצד השמאלי של שדה הראייה (משתי העיניים), וגרעין הברך הצדי בהמיספרה השמאלית מקבל מידע מהצד הימני של שדה הראייה.
תאי הגנגליון ברשתית מתחלקים למספר אוכלוסיות שונות של תאים שהן בעלות מורפולוגיה ייחודית. כל סוג של תא גנגליון מקושר לשכבה אחרת של תאי עצב בתוך גרעין הברך הצדי:
- תאי פאראסול (Parasol) מקושרים לשכבות בגרעין הברך הצדי שמכונות מאגנו-תאיות (Magnocellular). במסלול זה העיבוד מתמקד בזיהוי הבדלים בבהירות והוא רגיש מאוד בזיהוי תנועה. כל תא פאראסול מקבל מידע ממספר רב של תאים קולטי אור, והוא בעל שדה קלט גדול יחסית (ולפיכך הרזולוציה במסלול זה נמוכה יחסית).
- תאים ננסיים (Midget) מקושרים לשכבות הפארבו-תאיות (Parvocellular) של גרעין הברך הצדי, אשר מאפשרות הבחנה בפרטים דקים של הגירוי החזותי. במסלול זה ישנה תגובה משמעותית יחסית להבדלים בצבע, אך רגישות מוגבלת לתנועה ולבהירות. כל תא ננסי מקבל קלט ממספר מועט של תאים קולטי אור, ובמקרים רבים מתא קנה בודד.
- תאים דו-שכבתיים (Bistratified) מקושרים לשכבות הקוניו-תאיות (Koniocellular). תאים אלה הם בעלי שדות קלט גדולים מאוד, ובעלי רגישות בינונית לניגודיות. נכון להיום, תפקידם בתפיסה חזותית אינו ברור.
עיבוד בקליפת המוח
עריכה- ערך מורחב – קליפת הראייה
המידע מתאי העצב בשכבות השונות של התלמוס מועבר הלאה אל קליפת הראייה, אזור בקליפת המוח שמוקדש לעיבוד קלט ראייתי. העיבוד מתחיל באזור הראייה הראשוני, המכונה V1, אשר ממוקם בקצה האונה העורפית. משם, המידע ממשיך לאזורים נוספים שבהם מתבצע עיבוד חזותי מתקדם.
בתאי העצב של V1 ישנו מיפוי מלא של שדה הראייה, כאשר נקודות סמוכות על פני קליפת הראייה מייצגות נקודות סמוכות במרחב. עם זאת, המיפוי אינו פרופורציונלי – חלק גדול מ-V1 מוקדש לקלט שמגיע מהגומה המרכזית, למרות שמדובר בחלק קטן משדה הראייה.
לאחר העיבוד הבסיסי ב-V1, אזורים שונים לאורך מסלול העיבוד בקליפת המוח הראייתית מתמחים במאפיינים שונים של הגירוי החזותי: תפיסת צבע, תנועה, זיהוי אובייקטים וכו'. תאי העצב באזורים אלה הם בעלי שדות קלט גדולים יותר לעומת V1, והם מגיבים לטווח מצומצם יותר של גירויים.
רק כאשר המידע הראייתי מגיע לקליפת המוח אנו נעשים מודעים לגירוי החזותי. בהתאם לכך, אנשים עם פגיעה מוחית ב-V1 ידווחו על עצמם כעיוורים, גם אם העין והמסלולים העצביים האחרים לא נפגעו.
תאורה וצבע
עריכהרגישויות הקנים והמדוכים
עריכה
כל האופסינים מכילים קבוצה פונקציונלית בשם רטינל, המשנה את סידורה המרחבי כאשר היא בולעת פוטון בודד של אור. מידת הרגישות של הקולטן לתדרים אלקטרומגנטיים שונים מושפעת על ידי החלבון הקשור לרטינל. תאי קנים מכילים את הקולטן רודופסין (החלבון סקוטופסין מצומד לרטינל), שרגישותו לקרינה אלקטרומגנטית היא מרבית עבור פוטונים באורך גל 498 ננומטר. תאי מדוכים מכילים אופסינים ממשפחת הפוטופסינים, המורכבים אף הם מרטינל ומשייר חלבוני הנבדל בחומצות אמינו בודדות מהסקוטופסין המרכיב את קולטן הרודופסין. רשתיותיהם של מרבית בני האדם מכילות שלושה פוטופסינים שונים שרגישותם מרבית לאורכי הגל 420, 534 ו-564 ננומטרים. המדוכים המכילים פוטופסינים אלה מכונים מדוכים רגישי אורך-גל קצר (Short Wavelength Sensitive, בקיצור SWS או S), בינוני (M) וארוך (L). כפי שניתן ללמוד מן האיור, עקומי ההיענות של צבעני M ו-L (העקומים הירוק והאדום) דומים למדי. מקצת הנשים מבטאות סוג רביעי של צבען ברשתיותיהן, הנבדל מצבען L בחומצת אמינו יחידה. בגן המקודד פוטופסין רביעי זה התחלף הקודון TCT, המקודד לסרין, ב-GCT המקודד לאלנין. שינוי זה גורם להזחה צנועה של 3–4 ננומטרים בעקום ההיענות של הצבען הרביעי. בניסויים נמצא כי תפיסת הצבע של נשים אלו עשירה מזו של נשים וגברים אחרים.[8][9] גברים, מנגד, מועדים יותר ללקות בעיוורון צבעים היות שהם נושאים העתק יחיד של כרומוזום X, הנושא גנים המעורבים בביטוי פוטופסינים ברשתית.
לאור הדמיון התורשתי בין הפוטופסינים השונים לבין הרודופסין, היה זה אך טבעי לשער כי מקורם של הפוטופסינים במוטנטים של רודופסין שהתפלגו ממנו במהלך ההתפתחות האבולוציונית של בעלי חוליות. מנגד הציע גורדון וולס (Gordon Lynn Walls) בשנת 1942 כי תאי הקנים בעלי הרודופסין הם שהתפתחו מהמדוכים, לצורך שיפור רגישותם לאור קלוש.[10] השערה זו, שנשענה על מחקריו על רשתית דג הלאמפרי (Pouched lamprey, חסר לסתות ממשפחת הצמדיים), נתקלה בספקנות מצד הקהילה המדעית. [11] במפתיע, העלה חקר הקירבה התורשתית בין בעלי חוליות שונים ממצאים התומכים בהשערתו של וולס. נמצא כי עוד בעידן הקמבריון לפני 540 מיליון שנים בוטאו 4 פוטופסינים שונים בבעלי חוליות, בטרם התפצלו אלה לחולייתנים לסתניים ולחסרי לסתות. פוטופסינים קדומים אלה הסתעפו לשש קבוצות המכילות למעלה מאלף מינים של אופסינים, בהם הרודופסין המצוי בתאי קנים אנושיים[5]. בעלי החוליות של ימינו מבטאים חמישה גנים שונים של אופסינים ברשתיותיהם, המקודדים לקולטנים הפורשים את מקטע הספקטרום האלקטרומגנטי לו רגיש היצור[11]. חלקם מבטאים יותר מצבען אחד באותו תא קולט אור[11].
שלושת סוגי המדוכים ברשתית האנושית מפוזרים באופן אקראי לכאורה ברחבי הרשתית. מספר המדוכים מסוג M ו-L במרכז הרשתית משתנה באופן ניכר מאדם לאדם[9] וניתן לשער כי גם תפיסת הצבע משתנה מאדם לאדם. מכיוון שמדוכים שונים מגיבים במידה שונה לפוטונים באורך גל נתון, ניתן להבחין בין שטף האור לבין אורך הגל שלו על ידי השוואת תפוקותיהם של מדוכים מסוגים שונים.
ראיית יום וראיית לילה
עריכהתהליך הראייה מושתת על קליטת אור, ולכן ראייה בשעות הלילה ובסביבות חשוכות מצריכה רגישות גבוהה יותר של המערכת לפוטונים. בבעלי חיים אשר פעילים בעיקר בלילה, מערכת הראייה היא בדרך כלל רגישה יותר, אם כי הם מסתמכים גם על חושים אחרים שמאפשרים להם לתפקד בחושך.
בבני אדם, ההבדלים בין ראייה ביום או בסביבה עשירה באור לבין כזו המתבצעת בסביבה דלה באור נובעים מהמאפיינים של הקנים והמדוכים ברשתית.
הקנים הם רגישים יחסית לאור, ולכן מאפשרים לנו לראות בחשכה. הראייה שמתבצעת דרך הקנים מכונה ראייה סקוטופית. מפני שבקנים קיים רק סוג אחד של קולטן, ראייה זו אינה מאפשרת להבחין בין צבעים שונים. בנוסף, הראייה הסקוטופית אינה מתאפיינת ברזולוציה גבוהה, בשל המיעוט היחסי של תאי עצב שמעבדים את המידע שמגיע מהקנים. לעומת זאת, בסביבה מוארת מופעלת הראייה הפוטופית, שמתבססת בעיקר על המדוכים. מערכת זו היא פחות רגישה לאור, אך היא מאפשרת ראיית צבע והבחנה מדויקת יותר בפרטי הסביבה החזותית.
ההבחנה בין מאפייני הקנים והמדוכים באה לידי ביטוי גם באפקט פורקינייה – שינוי בתפיסת הצבע והניגודיות שמתרחש כתלות בכמות האור בסביבה. הקנים רגישים יותר לאור באורכי גל קצרים, ולכן בסביבה עם מעט אור עצמים כחולים ייראו בהירים יותר. בסביבה מוארת, הראייה נשלטת על ידי הרגישות של המדוכים, אשר מגיעה לשיא באורכי גל ארוכים יותר, ולכן עצמים אדומים למשל ייתפסו כבהירים יותר מסביבתם. תופעה זו נקראת על שמו של האנטומיסט הצ'כי שתיאר אותה, כאשר הבחין שהפרחים האדומים בגינתו נראים בהירים מאוד בשעות הצהריים, אך בשעות הדמדומים הם דווקא כהים ביחס לסביבתם. אפקט זה מתרחש רק בזמן המעבר בין ראיית יום לראיית לילה: בסביבה חשוכה מאוד המדוכים כמעט אינם פועלים, ולכן אין ראיית צבע.
הרפלקס האישוני
עריכה

- ערך מורחב – רפלקס אור
תפיסת הצבע האנושית
עריכההסתגלות לחשיכה ולאור
עריכההסתגלות לחשיכה
עריכההסתגלות לחשיכה (dark adaptation(אנ')) מערבת את הגדלת האישון בטווח הזמן הקצר, ותהליכים ביוכימיים לטווח הארוך. משך ההסתגלות הוא כחצי שעה[12][13].
תנועות עיניים
עריכה- ערך מורחב – תנועות עיניים

שישה שרירים אחראים על תנועות גלגל העין, והם מעוצבבים על ידי עצבים קרניאליים.
ראו גם
עריכהקישורים חיצוניים
עריכה- ארז גרטי, הראייה – אור ופעילות עצבית, במדור "מאגר המדע" באתר של מכון דוידסון לחינוך מדעי, 9 ספטמבר 2009
- יורם שורק, "ומתוק האור לעיניים", על חשיבות האור הנראה לבריאות, במדור "דברים שיורמים יודעים", באתר ארכיון האינטרנט (במקור, מאתר "nana10"), 13 במאי 2011
- אבי אליהו, בעיניים שלהם: איך בעלי חיים רואים את העולם?, באתר מאקו, 25 בפברואר 2015
- Sensation & Perception – סרטון הסבר על תפיסה וחישה עם התמקדות בחוש הראייה

- הרצאה של חמוטל סלובין על הקשר בין מערכת הראייה ועבודת המוח.
 גדעון לב, בניסוי בדגי זברה מצאו שפענוח גלי האור מתבצע כבר ברשתית, באתר הארץ, 3 בנובמבר 2021
גדעון לב, בניסוי בדגי זברה מצאו שפענוח גלי האור מתבצע כבר ברשתית, באתר הארץ, 3 בנובמבר 2021
הערות שוליים
עריכה- ^ 1 2 Lucas RJ, Hattar S, Takao M, Berson DM, Foster RG, Yau KW., Diminished pupillary light reflex at high irradiances in melanopsin-knockout mice, Science. 2003 Jan 10;299(5604):245-7. PMID 12522249
- ^ 1 2 Dacey DM, Liao HW, Peterson BB, Robinson FR, Smith VC, Pokorny J, Yau KW, Gamlin PD., Melanopsin-expressing ganglion cells in primate retina signal colour and irradiance and project to the LGN, Nature. 2005 Feb 17;433(7027):749-54. PMID 15716953
- ^ 1 2 Hattar S, Kumar M, Park A, Tong P, Tung J, Yau KW, Berson DM., Central projections of melanopsin-expressing retinal ganglion cells in the mouse, J Comp Neurol. 2006 Jul 20;497(3):326-49. PMID 16736474
- ^ ראו בקצרה תגובה לאור
- ^ 1 2 Akihisa Terakita, The opsins, Genome Biol. 2005;6(3):213 PMID 15774036
- ^ Nayak SK, Jegla T, Panda S., Role of a novel photopigment, melanopsin, in behavioral adaptation to light, Cell Mol Life Sci. 2007 Jan;64(2):144-54 PMID 17160354
- ^ Curcio CA & Allen KA (1990). "Topography of ganglion cells in human retina". The Journal of Comparative Neurology. 300 (1): 5–25. doi:10.1002/cne.903000103.
- ^ Jameson KA, Highnote SM, Wasserman LM., Richer color experience in observers with multiple photopigment opsin genes, Psychon Bull Rev. 2001 Jun;8(2):244-61., PMID 11495112
- ^ 1 2 Samir S Deeb, Genetics of variation in human color vision and the retinal cone mosaic, Curr Opin Genet Dev. 2006 Jun;16(3):301-7. PMID 16647849
- ^ Gordon Lynn Walls, The vertebrate eye and its adaptive radiation, Cranbrook Press, Bloomfield Hills, Michigan, 1942, 14 + 785 pp.
- ^ 1 2 3 Collin SP, Trezise AE, The origins of colour vision in vertebrates, Clin Exp Optom. 2004 Jul;87(4-5):217-23. PMID 15312025
- ^ https://www.scientificamerican.com/article/experts-eyes-adjust-to-darkness/
- ^ נטע סופר צור, למה הסתגלות העיניים לחושך נמשכת כל כך הרבה זמן?, באתר הארץ, 26 במרץ 2019