מערכת השמיעה
מערכת השמיעה היא מערכת החישה, האחראית על חוש השמיעה ומאפשרת קליטת קולות. המערכת אחראית על התמרת הקלט הפיזיקלי השמיעתי אל אות חשמלי והעברתו למרכזי העיבוד המרכזיים במוח, על ניתוח מרכיביו והפקת מידע מכך ועל זיהוי מקורו. לעיתים לשם פעולות הניתוח נעזרת מערכת השמע במידע המגיע ממערכות חוש אחרות כגון מערכת הראיה, וכן בניבויים אשר הוכנו על בסיס ידע קדום מבעוד מועד[1].
_(20374669702).jpg) | |
| שיוך |
מערכות חישה |
|---|---|
הגירוי השמיעתי עריכה

 ערך מורחב – קול
ערך מורחב – קול
הגירוי אותו מערכת השמע מנתחת על מנת להפיק מידע הוא גלי קול. האוויר העשוי מחלקיקים שונים יכול להידחס ולהתרווח כתלות בגורמים פיזיקליים שונים כמו טמפרטורה. לעיתים קרובות דחיסות האוויר מופיעה במחזוריות מסוימת, אשר ניתנת לאפיון כגל ונהוג לקרוא לו גל קול. מערכת השמע בנויה כך שהיא מתמירה את תכונות הגלים הפיזיקליות משרעת, תדר ורמת מורכבות אל עבר הממד התפיסתי הכולל את עוצמת הצליל (Loudness), גובהו (Pitch) וגוונו (Timbre) בהתאמה[2].
אנטומיה של האוזן עריכה
- ערך מורחב – אוזן
האוזן מחולקת לשלושה חלקים נפרדים: האוזן החיצונית, האוזן התיכונה והאוזן הפנימית[2].
האוזן החיצונית עריכה
החלק הנראה לעין של האוזן נקרא אֲפַרכֶּסֶת, הוא עשוי מסחוס וקיים רק ביונקים. עיצוב האפרכסת מותאם לקליטת קולות ולכוונם אל תוך חלקי האוזן הפנימית, ובכך לשפר את יכולת קליטת הקול מהסביבה. האפרכסת מתעלת את גלי הקול לתוך תעלת השֶמַע, שבקצה נמצא עור התוף. תעלת האוזן בחלקה החיצוני עשויה מסחוס, ובעומקה, כשני שלישים ממנה נמצא בתוך עצם. היא מצופה בעור דק, המשכי לעור האפרכסת. בקצה התעלה העמוק נמצא עור התוף, הוא מתוח בקצה צינור השמע, ומעברו השני האוזן התיכונה[2].
האוזן התיכונה עריכה

האוזן התיכונה נמצאת בין עור התוף אל השבלול במרווח בנפח של שני מיליליטר. מבחינה תפקודית האוזן התיכונה משמשת כמגבר המגביר את תנודות עור התוף ומעביר אותן לאוזן הפנימית המלאה בנוזל. התנודות עוברות לאורך שלוש עצמות קטנטנות. העצם הראשונה, נקראת עצם הפטיש (Malleus), מחוברת לעור התוף ונעה כאשר הוא רועד. כאשר עצם הפטיש נעה היא מזיזה את העצם השנייה – עצם הסַדָן (Incus), אשר בתורה מניעה את העצם השלישית – עצם הארכוף (Stapes). התנודות עוברות דרך הארכוף אל החלון הסגלגל, שהוא למעשה קרום הנמצא בכניסה אל האוזן הפנימית.
עצמות השמע מחד, והפרש השטחים בין שטח עור התוף לשטח משטח הדריכה של הארכוף, גורמים להגברה של הכוח שמפעיל גל הקול פי שלושים ובכך הם תורמים להעברה יעילה של הקול מן האוויר לתנודות של גלי נוזל באוזן הפנימית. מלבד יכולת ההגברה, לאוזן התיכונה תפקיד חשוב בשמירה על השבלול מפני גלי קול חזקים מדי שעלולים לגרום נזק לשמיעה. הארכוף ניחנת ביכולת לשנות את זווית ההקשה, כך שבעת הצורך הכוח המועבר לחלון הסגלגל מופחת. כמו כן, ישנם שרירים קטנים אשר גלים בתדר נמוך ובעוצמה גבוהה מפעילים אותם במנגנון רפלקסיבי היוצר מתיחה של עור התוף, ונעילה של הארכוף[3].
האוזן הפנימית עריכה
האוזן הפנימית חצובה בגולגולת והיא נמצאת פנימה לעצם הארכוף. באוזן הפנימית מצוי מבנה מסולסל הנקרא שבלול האוזן יחד עם מבנה אחר האחראי על שיווי המשקל. שבלול האוזן מלא נוזל בעל תכונות כימיות מיוחדות השונות מהנוזל החוץ תאי בשאר רקמות הגוף. לשבלול שלושה מדורים המהווים יחד שלוש תעלות שרצות בצורת ספירלה: מַעֲלַת הַפְּרוֹזְדוֹר "scala vestibuli", מעלת האמצע "scala media" ומַעֲלַת הַתֹּף "scala timpany". במעלת התוף ומעלת הפרוזדור נע נוזל בשם פרילימפה, ואילו במעלת האמצע נע נוזל בשם אנדולימפה.
כאשר עצם הארכוף רוטטת היא פוגעת בחלון הסגלגל ודוחפת את הנוזל שבשבלול. הנוזל נע ראשית במעלת הפרוזדור, מתקדם אל עבר מעלת התוף, ושם מרטיט את החלון העגול. נהוג להתייחס לשבלול כבעל ציר בעל שני קטבים שרץ לאורך הספירלה. קוטבו הבזאלי (הבסיס) מתחיל בחלון הסגלגל, וקוטבו האפיקלי (הקודקוד) נמצא בקצה השבלול. לו היו פורמים את גלגול השבלול לצינור עם שלוש תעלות פנימיות, היה הצד האפיקלי קצה הצינור שנוצר מהפרימה. בין מעלת התוף לבין מעלת האמצע ישנו קרום הנקרא הקרום הבזילרי, ובין מעלת הפרוזדור למעלת האמצע נמצא קרום רייזנר המחובר לקרום הטקטוריאלי. כאשר הנוזל נע בתעלות הפרוזדור והתוף שבשבלול הוא מעוות את הקרום הבזילרי בקצב מסוים ובמקומות ספציפיים. בין הקרומים נמצא איבר קורטי המשמש כיחידה בה מותמר הקלט השמיעתי לעבר אות חשמלי[2].
איבר קורטי עריכה

בתוך מעלת האמצע בשבלול האוזן נמצא איבר התמרת השמע, איבר קורטי (אנ'). איבר קורטי מורכב מהקרום הבזילרי, מהקרום הטקטוריאלי, מתאי דייטר המתפקדים כתאי תמך ומתאי שערה המתפקדים כתאי חישה. תאי חישה אלו הם בעלי שלוחות הקרויות "ריסים (cilium)" העשויות מאקטין ומיוזין[4] היוצאות מגוף התא הדומות לשערות קטנות ומכאן שמו. לתאים אלו סינפסות עם שלוחות של תאי גנגליון דו–קוטביים השייכים לגנגליון בשם הגנגליון הספירלי.
התנודות של מרכיבי האוזן התיכונה מועברות אל הנוזל שמקיף את איבר קורטי, וגורמות להזזת הממברנה הטקטוריאלית ביחס לממברנה הבזילרית. תזוזה זו מזיזה את השערות הנמצאות על תא השערה. שערות אלו מחוברות זו לזו במין חוטים הנקראים Tip Link המחוברים אל תעלות יוניות[4]. כאשר ישנה תזוזה בין תאי השערה ה-Tip Link נמתח ובכך הוא גורם לפתיחה מכנית של התעלות היוניות[4]. פתיחת התעלות היוניות גורמות לכניסה של יוני אשלגן ויוני סידן[5] אשר נמצאים בריכוז גבוה בסביבה החוץ תאית של השבלול ובכך משנות את פוטנציאל הממברנה של התאים. שינוי פוטנציאל הממברנה גורם לשחרור וסיקולות אל קולטנים בתאים הביפולארים של הגנגליון הספיראלי[2].
-
 חתך אורכי של השבלול המראה הן את המדורים השונים והן את מיקום איבר קורטי.
חתך אורכי של השבלול המראה הן את המדורים השונים והן את מיקום איבר קורטי.

ישנם שני סוגי תאי שערה, בין 12,000 ל-15,000 תאי שערה חיצוניים וכ-3,500 תאי שערה פנימיים[6]. תאי השערה הפנימיים הם אלו אשר מעבירים את רוב המידע החושי אל מערכת העצבים המרכזית. לעומתם תאי השערה החיצוניים באמצעות תאי דייטר משנים את האופן בו הממברנה הבזילרית זזה[7][8]. בכך הם משפיעים על המידע הנקלט בתאי השערה הפנימיים, ועוזרים בהגברת קלט חלש[3].
התמרה חושית של הגירוי השמיעתי עריכה
שינויים בקלט החושי משנים את מקדם החדירות לתעלות יוניות הנמצאות על הקצה החיצוני של תאי השערה ובהתאמה לכך את דפוס הדחפים העצביים המועברים אל מערכת העצבים המרכזית. ככל שהמוליכות יותר גבוהה כך קצב ירי פוטנציאלי הפעולה בתאים הדי פולאריים עולה[2].
קידוד תדר הצליל עריכה
_he.svg)
זיהוי התדר של צלילים מתבצע עוד בשבלול ומתחלק לשני אופנים מרכזיים: על ידי זמן, ועל ידי מקום.
קידוד על פי תדירות הרעידות של הקצה הקודקודי עריכה
מתחת ל-200 הרץ הקרום הבזילרי אינו מפגין הבחנה בין תדרים שונים על פי מיקום הרעידה המקסימלית[9]. במקרים אלו ההבחנה בין תדרים שונים נעשית על פי תדירות הרעידות על הקצה הקודקודי (אפיקלי). ככל שהרעידות הן בתדר גבוה יותר, כך הצליל הנתפס הוא יותר גבוה[10][11].
קידוד על פי מרחק מהחלון הסגלגל עריכה
שוני בגמישות הקרום הבזילרי וברוחבו לאורך השבלול מסייע לפרק את צלילי הסביבה לתדרים: ככל שמתרחקים מהצד הבסיסי (בזאלי) של השבלול אל עבר קודקוד השבלול, כך התדר בו הממברנה תרעד בצורה מקסימלית נמוך יותר. ככל שהרעד רב יותר, כך נפתחות יותר תעלות יוניות, משתחרר יותר מוליך עצבי, ותדר פוטנציאלי הפעולה שיורה הנוירון הבי-פולארי עולה[2].
קידוד עוצמת הצליל עריכה
בתדרים היורדים עד ל-200 הרץ עוצמת הצליל מזוהה על ידי תדירות הירי. הרעדה חזקה יותר של עור התוף ועצמות השמע גוררת הרעדה חזקה יותר של הקרום הבזילרי, שבתורו מכופף את תאי השערה ופותח יותר את התעלות היוניות. פתיחת התעלות גורמות לשחרור מוגבר של מוליכים עצביים, אשר בתורם מגבירים את קצב הירי בתאי העצב הספירליים. כאשר התדרים יורדים מתחת ל-200 הרץ, קידוד עוצמת הקול מתבצע על ידי כמות הנוירונים היורים בו זמנית. ככל שהעוצמה חזקה יותר כך יותר נוירונים יורים[2]. יחד עם הסבר זה ישנם גורמים נוספים המשפיעים על העוצמה הנתפשת של הגירוי המושמע כדוגמת משך זמן ההשמעה, והאם הוא נשמע בשתי האוזניים או באוזן אחת[3]. נראה כי בטווח הזמנים של עד 200 מילישניות ככל שזמן השמעת הצליל גדול יותר כך גדלה עוצמת הצליל הנתפסת. כמו כן, כאשר הצליל מושמע בשתי האוזנים העוצמה נתפשת כגדולה יותר מאשר אותו הגירוי המושמע לאוזן אחת[3].
קידוד מיקום הצליל עריכה
תפיסת מיקום הצליל ביחס למיקום האורגניזם היא משימה מורכבת ומתרחשת, לפחות בחלקה, בגזע המוח בגרעינים הנקראים MSO ו-LSO. מבחינת המידע המשמש לזיהוי מקור הצליל ידוע כי נעשה שימוש בהפרשי פאזות של הגל הנקלט בין שתי האוזניים, בהפרשי עוצמה מזעריים בין שתי האוזניים ובהשתנות גלי הקול בעת פגיעתם באפרכסת.
מחקרים שמדדו את הזווית הקריטית המינימלית שבה בני אדם וקופים תופסים צלילם מצאו שהיא עומדת על מעלה אחת[12] או על ארבע מעלות[13] בהתאמה כאשר מופנה קשב למשימה.
מעגלי המוח עריכה
בעיבוד מקור הצליל מעורבים שני גרעינים בגזע המוח הנקראים הגרעין הזיתי האמצעי-עליון (MSO) והגרעין הזיתי הצידי-עליון (LSO). התאים ב-MSO מקבלים מקבלים את המידע כאות מאקטב משתי האוזניים, ומתבצע בו חישוב של הפרש הזמנים של האות הנקלט באוזן אחת אל מול האוזן השנייה. בתא הנמצא באמצע, בין צד ימין לשמאל ככל שההפרש של הזמנים של האות המגיע מהאוזניים יותר קטן כך התא יירה יותר. תאים הקרובים יותר לצד מסוים יירו הכי הרבה אם יש הפרש זמנים בין האותות. לעומת זאת ב-LSO הקלט הוא מצד אחד מאקטב ומצד אחר מחליש. בגרעין זה מבוצע חישוב של הפרשי העוצמות בין האוזניים.
-
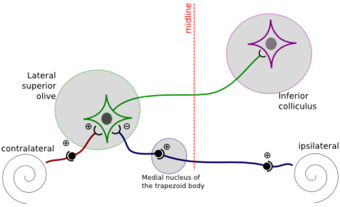 מעגל עיבוד מיקום הצליל בגרעין ה-LSO
מעגל עיבוד מיקום הצליל בגרעין ה-LSO -
 מעגל עיבוד מיקום הצליל בגרעין MSO
מעגל עיבוד מיקום הצליל בגרעין MSO


קידוד גובה הצליל עריכה
גובה-הצליל (pitch) הוא גובה הצליל הנתפס בדעת, אשר מתבסס על זיהוי התדר היסודי של גל הקול שנקלט במערכת השמע. תדר זה מוגדר כהופכי לזמן המחזור של גל הקול שנקלט[14].
גל ניתן לפירוק, על ידי שיטה מתמטית בשם התמרת פורייה, לסדרה של גלים סינוסידיאלים. התדרים של גלים אלו אשר שונים מתדירות בעלת מאפיינים פיזיקליים ייחודיים הנקראת תדירות יסודית נקראים תדרים עילאיים. התדרים העילאיים הם אלו אשר מעניקים את הגון (timber) לצליל, ומפרידים בין צלילים בעלי אותו גובה המושמעים מכלי נגינה שונים. פירוק דומה להתמרת פורייה נעשה בצורה מכנית בתוך השבלול על פי מרחק ועל פי פאזה כפי שמתואר לעיל בפסקת "קידוד תדר הצליל". כאשר גל הקול שנקלט אינו מכיל את התדר היסודי, מערכת השמיעה יודעת לפענח על סמך התדרים העילאיים את התדר היסודי שיכול היה להיות, והצליל שייתפס הוא הצליל המתאים לתדר יסודי זה. יכולת זו נצפתה אף אם התדרים העילאיים המושמעים שונים בכל אחת מהאוזניים, ומתמטית חילוץ התדר היסודי נדרש על ידי שילוב מידע של שתי האוזניים[3]. בנוסף לזאת נמצא כי היכולת לתפוס את גובה הצליל, אף כשאין בנמצא תדר יסודי, אינה ייחודית לבני אדם, והיא נצפתה גם אצל חיות כמו קופים[15], ציפורים[16] וחתולים[17].
המסלול העצבי של השמע עריכה
לאחר ההתמרה בשבלול הקלט החושי עובר אל עצב השמיעה ושיווי המשקל (CN VIII) דרך הגנגליון הספירלי[2]. עצב השמיעה מכיל אוכלוסייה הטרוגנית של תאים, המציגים תכונות חישתיות שונות. חלק מהתאים מגיבים רק לצלילים מורכבים, אחרים רק לטהורים. בנוסף יש כאלה אשר מגיבים רק לצלילים מאוד קצרים או לחלופין מגיבים לקשת גדולה של צלילים מסוגים שונים. אוכלוסיות תאים אחת השוכנת בעצב השמיעה היא של תאים מכוונים. תאים אלה מגיבים למנעד של תדירויות, אך במיוחד לתדר ספציפי. כלומר, סף התגובה לירי פוטנציאל פעולה בתדר ספציפי זה יהיה נמוך מסף התגובה לירי פוטנציאל פעולה בכל תדר אחר עבור אותם תאים. עם זאת אין הדבר אומר כי תדרים אחרים אינם יכולים להביא לירי. אלא צריך בתדרים אלה גירוי בעוצמה גדולה יותר כדי להגיע לאותו ירי המופק מגירוי חלש בתדר הספציפי. בעיקרו מקור סלקטיביות זו מצוי בסלקטיביות הממברנה הבזילרית לתדרים שונים במקומות שונים. אך במחקרים נמצא כי אוכלוסיית תאי עצב השמיעה המכוונים מציגים סלקטיביות גבוהה מזו המסופקת באמצעות הממברנה הבזילרית. דבר המוסבר על ידי שינויים אקטיביים של רכיבים נוספים באיבר קורטי[3].
מהגנגליון הספירלי מועבר המידע אל מספר גרעינים במוח המוארך. במוח המוארך מתבצע עיבוד בסיסי של הצליל. לדוגמה זיהוי גס של הכיוון ממנו הגיע השמע. המידע מועבר ראשית לגרעין השבלול הגחוני (Ventral cochlear nucleus (אנ')) ולגרעין השבלול הגבי (Dorsal cochlear nucleus (אנ')). מהם עובר הקלט אל קומפלקס של מספר גרעינים הנקרא הגוש הזיתי העליון (Superior olivary complex (אנ')) ואל הגוף דמוי טרפז (Trapezoid body (אנ')). ולאחריהם עובר המידע באמצעות צבר אקסונים הנקרא הלמניסקוס הצידי (Lateral lemniscus (אנ')) אל התליל התחתון שבמוח האמצעי. ממנו מועבר המידע אל גרעין הברך האמצעי (Medial geniculate nucleus (אנ')) שבתלמוס ולקליפת המוח השמיעתית ראשונית (Primary auditory cortex (אנ'))[2]. מלבד מסלולים אלו, ההולכים אל מערכת העצבים המרכזית מהמערכת העצבים הפריפריאלית, ישנם גם מסלולים הפוכים בכיוונם, המכונים מסלולים יורדים או מוציאים (אפרנטים – Efferent pathways), לאורך כל מערכת השמע[18]. מסלולים אלו מאפשרים לאזורים גבוהים בקליפת המוח לווסת את תהליכי עיבוד המידע במערכת העצבים הפריפריאלית[19][20].

קליפת המוח השמיעתית עריכה
חלוקה היררכית עריכה

קליפת המוח השמיעתית מחולקת בגסות לשלושה אזורים היררכים בעיבוד המידע השמיעתי: אזור הליבה (Core region), אזור הרצועה (Belt region) והאזור שליד הרצועה (Parabelt region)[2]. האזור השמיעתי הראשוני חבוי בתוך המענית הצידית על האזור המדיאלי בפיתול על שם השל[22]. הוא בנוי על פי קידוד התדרים המרחבי שבשבלול – ככל שמתקדמים לכיוון יותר קדמי (Anterior) כך התדר שיגרום לתגובה המקסימלית של תאי העצב באזור נמוך יותר. לצורת ארגון מסודרת זו, של התגובה לתדרים שונים, קוראים ייצוג טונוטופי. הייצוג הטונוטופי מופיע עוד פעמים לאורך קליפת המוח השמיעתית[21], כפי שאפשר לראות בתמונה משמאל. אזור מרכזי נוסף במערכת השמע, שנמצא בקליפת המוח, הוא מרכז הגובה (Pitch center). מרכז זה נמצא לטרלית אל הקורטקס האודיטורי הראשוני, והוא מעורב רבות בקידוד הגובה הנתפס של הצליל (Pitch)[22].
מסלול ה"מה?" ומסלול ה"איפה?" עריכה
בדומה לארגון של אזורים ראייתים גם בקליפת המוח השמיעתית האסוציאטיבית ישנם שני ערוצי עיבוד מידע מובחנים: ערוץ קדמי וערוץ אחורי. הערוץ הקדמי אשר נקרא גם מסלול ה"מה?" מתחיל בחלק הקדמי של האזור שלצד הרצועה, והוא עוסק באנליזה של הקלט השמיעתי. הערוץ האחורי אשר נקרא גם מסלול ה"איפה?" מתחיל בחלקים אחוריים של האזור שליד הרצועה, והוא מתמקד באיכון מקור הצליל. ההפרדה בין המיקומים נצפתה אף ברמת הנוירון הבודד[23] וגם בניסויי דימות מוחי בבני אדם[24]. כמו כן נצפתה גם סיבתיות, פגיעה באחד מאזורים אלו שיבשה את היכולת לפענח מידע של מיקום או של זיהוי בהתאמה למיקום הפגיעה[25].

גמישות תפקודית עריכה
תכונה של קליפת המוח היא היכולת שלו להפגין קישוריות דינאמית בין האזורים שונים. תכונה זו באה לידי ביטוי בקליפת המוח השמיעתית בכמה אופנים. לדוגמה: כאשר יש שימוש מוגבר בחוש השמיעה מתרחשים שינויים מבניים. אצל עיוורים הקורטקס השמיעתי נמצא מקושר יותר לקורטקס הראייתי. בבחינה תפקודית נמצא שאצל העיוורים בהינתן קלט שמיעתי, הקורטקס הראייתי שלהם הגיב ביתר שאת, דבר המעיד על גלישה של עיבוד השמע אל עבר אזורים אלו[27]. דוגמה נוספת היא מקליפת המוח השמיעתית של מוזיקאים. נמצא כי אצל מוזיקאים הפיתול על שם השל גדול יותר, ופעיל יותר. יתרה מכך, הן הגדילה בנפח והן הפעילות המוגברת נמצאו ביחס ישר לכישורים המוזיקליים של הנגן[28].
תפקודים גבוהים במערכת השמע – שמיעה מוזיקלית הבנת שפה ומנגנוני ניבוי עריכה
תפיסה מוזיקלית במערכת השמע היא תהליך מורכב. מלבד פירוק הצלילים לתכונותיהם הבסיסיות המערכת השמיעתית מסוגלת גם למצוא תבניות ברצפי צלילים. לפי תבניות אלו המערכת מסוגלת לנבא צלילים העתידים לבוא[29]. בפעילות זו מעורבים אזורים מוחיים גבוהים, המתפרשים על קליפת המוח הקדם מצחית ועל קליפת המוח הרקתית[30]. היכולת לנבא את הצלילים הבאים מאפשרת לזהות רצפי צלילים בעלי משמעות גבוהה במיוחד. זאת לאור עקרון בסיסי בתורת האינפורמציה, שלפיו רצף הניתן לניבוי בצורה מלאה אינו נושא עמו אינפורמציה חדשה[31]. על כן, מערכת מוחית אשר יודעת לנבא צלילים יכולה למקד את משאביה לטובת עיבוד צלילים מפתיעים, המכילים מידע רב. היא מבצעת זאת באמצעות מנגנונים כמו הפניית קשב[32].
לאורך מערכת השמיעה קיימת רשת עצבית האחראית על איתור רצפי קול מפתיעים. רצפים שחורגים ממה שמצפה המערכת העצבית[33]. בין תהליכי האיתור אופיין במערכת העצבית של האדם גל בשם Mismatch negativity או בקיצור MMN[34]. ה-MMN מופיע לאחר הופעת קלט מפתיע, קלט הסותר ניבוי שנוצר ברשת העצבית לגבי העתיד הקרוב[35][36]. גל זה מופיע בדמות פוטנציאל קשור-אירוע (ERP), דהיינו התפלגות פוטנציאל חשמלי ייחודי על הקרקפת, הניתן למדידה באמצעות מכשיר EEG[37]. האזורים המעורבים ביצירת גל זה מאותתים כי היה קלט מפתיע. לכן, באמצעות זיהוי הגל ניתן להסיק כי המוח "הופתע". האזורים שיוצרים את הגל הם הפיתול המצחי התחתון, והפיתול על שם השל. איתור רצפי קול מפתיעים, המעוררים את גל ה-MMN, הוא תהליך שאינו דורש קשב ולרוב גם לא מודעות. התהליך נצפה גם במטופלים שהיו שרויים בתרדמת[38].
-
![דוגמה לשינויי פוטנציאל הנגרמים בתופעת ה-Mismatch negativity. ניתן לראות את הפוטנציאל קשור-אירוע האופייני בקו האדום בתמונה A. בתמונה B מופיע תרשים המתאר בצבעי כחול-אדום את השתנות הפוטנציאל החשמלי כפונקציה של הזמן (ציר אופקי) באלקטרודות שונות של ה-EEG (מפוזרות על הציר האנכי)[39].](//upload.wikimedia.org/wikipedia/commons/thumb/6/6b/Fnins-07-00265-g001.jpg/500px-Fnins-07-00265-g001.jpg) דוגמה לשינויי פוטנציאל הנגרמים בתופעת ה-Mismatch negativity. ניתן לראות את הפוטנציאל קשור-אירוע האופייני בקו האדום בתמונה A. בתמונה B מופיע תרשים המתאר בצבעי כחול-אדום את השתנות הפוטנציאל החשמלי כפונקציה של הזמן (ציר אופקי) באלקטרודות שונות של ה-EEG (מפוזרות על הציר האנכי)[39].
דוגמה לשינויי פוטנציאל הנגרמים בתופעת ה-Mismatch negativity. ניתן לראות את הפוטנציאל קשור-אירוע האופייני בקו האדום בתמונה A. בתמונה B מופיע תרשים המתאר בצבעי כחול-אדום את השתנות הפוטנציאל החשמלי כפונקציה של הזמן (ציר אופקי) באלקטרודות שונות של ה-EEG (מפוזרות על הציר האנכי)[39].
![דוגמה לשינויי פוטנציאל הנגרמים בתופעת ה-Mismatch negativity. ניתן לראות את הפוטנציאל קשור-אירוע האופייני בקו האדום בתמונה A. בתמונה B מופיע תרשים המתאר בצבעי כחול-אדום את השתנות הפוטנציאל החשמלי כפונקציה של הזמן (ציר אופקי) באלקטרודות שונות של ה-EEG (מפוזרות על הציר האנכי)[39].](/wiki/%D7%A7%D7%95%D7%91%D7%A5:Fnins-07-00265-g001.jpg)
רישומי מוח שפורסמו בשנת 2009 מראים כי לא רק שיש אזורים שמתריעים "יש הפתעה בקלט", אלא גם שאם ההפתעה חוזרת שוב ושוב – בצורה הניתנת לחיזוי – אז עם הזמן המוח יתחיל לצפות לה. במקרה שכזה נאמר כי התהוותה "ציפייה מסדר גבוה" (המוח מצפה להפתעה, כלומר לחריגה מהמצופה)[38]. עם זאת, ציפייה מסדר גבוה כבר דורשת פניות של המיינד. רישומי EEG, שנלקחו מנבדקים שנדרשו לעשות מטלה הדורשת מאמץ מחשבתי רב, כמעט ולא הראו סממנים לציפייה מסדר גבוה[38]. ממצא מעניין הוא, שמטופלים בתרדמת שהציגו ציפייה מסדר גבוה, יצאו מהתרדמת תוך כמה שבועות. מטופלים שלא הציגו יכולת זו נשארו בתרדמת יותר זמן או לא יצאו ממנה כלל[38]. רישומי המוח שפורסמו בשנת 2009 הראו עקבות לציפייה שכזו, בשנת 2019 חוקרים הצליחו לזהות סממן ישיר לציפייה מסדר גבוה: גלים מסוימים בתדר גבוה, שנוצרים באונה הפרונטלית[40].
הנאה ממוזיקה וחוויית צמרמורת עריכה

אזורים גבוהים במסלול השמע מתקשרים עם מערכת התגמול ומובילים לחוויית ההנאה ממוזיקה. חוויית הנאה זו יכולה להגיע לשיא ולהוביל לצמרמורת משולבת בפרץ גדול של הנאה ובשינוי ניכר בדפוס המוליכות העורית וקצב פעימות הלב[41]. נמצא כי רצפי צלילים אשר מובילים לצמרמורת מאופיינים לרוב בחידוש כלשהו באחד מממדי המוזיקה, כמו עליה פתאומית בגובה הצלילים, הגברת עוצמת הקול, החלפת כלי הנגינה הראשי או נגינה של התזמורת שהייתה לא מתואמת נהפכה פתאום מתואמת[42].
הבנת שפה עריכה
בתחילת המחקר המדעי של הבנה והפקת שפה התגלו שני אזורים מרכזיים המעורבים בתהליך מלבד הקליפה השמיעתית ראשונית. הראשון נקרא אזור ברוקה והשני נקרא אזור ורניקה. עם השתכללות כלי המחקר המדעי והתקבלות הגישה הדו-מסלולית כגישה מרכזית ומתומכת עודכנו המודלים, וכיום יודעים על תתי אזורים המתמחים בעיבודים שונים של השפה[43]. בנוסף להבנה מבנית ותפקודית טובה יותר של האזורים השפתיים, עם הזמן נחשפו מנגנונים גבוהים בקליפת המוח העוזרים בעיבוד השפה. לדוגמה: נמצא כי תנודות חשמליות הנוצרות בקליפה השמיעתית נמצאות בהתאמה למקצבים האופייניים לדיבור אנושי, אשר מעורבות בהבנת הדיבור[44]. בנוסף, נמצא כי מנגנונים קשביים יכולים לווסת את תנודות אלו, ובכך הם מאפשרים התמקדות בדובר מסוים על פני אחרים[45][46]. כמו כן, בקליפת המוח מתבצעת אינטגרציה של מידע מחושים רבים, תהליך העוזר גם הוא לפענוח הדיבור[47].
מחלות המקושרות למערכת השמיעה עריכה

הפרעות שונות נגרמות בשל כשלים במקומות שונים במערכת השמיעה. חלק מהבעיות הן התפתחותיות וכוללות עיוותים בגדילת האוזן או מיקרוטיה. בעיות אחרות נובעות מטראומה כמו נקב בעור התוף, הסרה חלקית או מלאה של האפרכסת, אוזן כרובית, חדירת גוף זר וכדומה. בנוסף לכך ישנן מחלות נרכשות של מערכת השמיעה. ישנם זיהומים חיידקיים, ויראלים או פטרייתיים כמו זיהום האפרכסת ((אנ') Perichondritis), דלקת האוזן החיצונית, דלקת פטרייתית של תעלת האוזן החיצונית (Otomycosis), דלקת האוזן התיכונה (Otitis Media), סיבוך נדיר של שלבקת חוגרת בשם תסמונת רמזי-האנט, כולסטאתומה, טרשת האוזן[48].
בנוסף, ייתכן גם כשל עצבי שמקורו במוח הגורם לבעיות שמיעתיות. לדוגמה חוסר היכולת לשמוע מוזיקה[24] (אמוזיה) או היווצרות הזיות שמיעתיות של מוזיקה[49] או של צלילים מטרידים (טנטון)[50].
בדיקת הולכה עריכה
לעיתים קרובות על מנת לאבחן בעיה במערכת השמע משתמשים בקולן, כלי מתכתי קטן המשמיע צליל בתדר אחיד לאחר הקשה עליו. עם הזמן עוצמת הצליל הבוקעת מהקולן הולכת ופוחתת. הצליל הנוצר על ידי הקולן מועבר על ידי האוויר למערכת השמע, אך אם הבודק יצמיד את הקולן לראש של הנבדק אזי הצליל הנוצר מרעד הקולן יעבור דרך העצם אל מערכת השמע. לרוב ההצמדה תעשה לעצם המסטואיד. בדיקה המבוססת על עובדות אלו נקראת מבחן רינה (Rinne Test). במצב בריא נבדקים שומעים יותר טוב צלילים הנישאים על תווך של האוויר. אם הנבדק מפסיק לשמוע את צליל הקולן, אך בהצמדה של הקולן לעצם הוא שב לשמוע את הצליל, הדבר לרוב יעיד על בעיה הולכתית. כלומר לבעיה בהולכת הצלילים אל מערכת העצבים.
בדיקה נוספת היא הצמדת הקולן לקו האמצע של הראש, על חלקו העליון. הבודק יברר אם קיימת א-סימטריה בין האוזניים של עוצמת הצליל. כאשר הבעיה הולכתית, הצליל יהיה יותר חזק בצד הפגוע, אך אם הבעיה היא עצבית אז הצליל יהיה חלש יותר בצד הפגוע. לבדיקה זו קוראים "מבחן ובר" (Weber Test)[51].
תהליכי הזדקנות במערכת השמיעה עריכה

הזדקנות של מערכת השמיעה המובילה לאיבוד יכולות שמיעתיות (Presbycusis) הוא תהליך מאוד נפוץ, הפוגע ב-80% מהמבוגרים מעל גיל 70[53]. לעיתים מתרחשת גם פגיעה חבויה, כזאת אשר אינה ניתנת למדידה ישירה באמצעות בדיקה אודיומטרית. פגיעות כאלו כוללות למשל הפחתה ביכולת להקשיב לקלטים שמיעתיים[54]. לתהליכי איבוד השמע בעקבות הזדקנות נמצאו גם השפעות נלוות, הכוללות בידוד חברתי ודיכאון, לחץ ועייפות נפשית (Mental fatigue)[55].
שינויים עצביים לאורך מערכת השמיעה עריכה
ההזדקנות משפיעה על כל שלבי השמיעה: מהתמרת האות בשבלול, הולכתו דרך גרעינים בגזע המוח ועיבודו הסופי בקליפת המוח[56]. אחת הפגיעות הנפוצות היא הידלדלות אוכלוסיית תאי השערה החיצוניים, בעיקר בקצה הקודקודי של השבלול. תאי שערה אלו מעורבים בעיקר בהגברת צלילים גבוהים, דבר המסביר את התופעה הנפוצה בזקנה של לקות שמיעה הממוקדת לצלילים בעלי תדר גבוה[6], המתבטאת בין היתר בצורך בצלילים חזקים יותר על מנת להגיע לסף השמיעה[57]. מחקרים בבעלי חיים מצאו כי מלבד הידלדלות אוכלוסיית תאי השערה החיצוניים, ישנו גם ניוון של האקסונים של עצב השמיעה, ותת פעילות של הסינפסות בעצב זה[58][59][60]. מעבר לפגיעות אלו נצפו בחיות ובבני אדם גם פגיעות בתאי עצב הגנגליון הספירלי[61], בגרעיני השבלול[62], בגוש הזיתי העליון, ובאזורים נוספים במעלה גזע המוח עד לתליל התחתון[63][64]. לרוב פגיעות בגרעיני גזע המוח ישפיעו על יכולות עיבוד מרווחי הזמן בקלטי שמע[65][66][67], ועל היכולת להבין דיבור[68][69][70].
אחד הסמנים הנפוצים לתהליכי ההזדקנות של המערכת שנצפו באדם[71] ובבעלי חיים[72] הוא שינוי בדפוס תגובות חשמליות בגזע המוח לגירוי שמיעתי (Auditory Brainstem Response – ABR), שינוי זה נמדד באמצעות מכשיר EEG. ברמה המוחית בבעלי חיים[73][74] ובבני אדם[75][76] נמצאה פעילות פחותה של מערכת ה-GABA בקליפת המוח. ה-GABA הוא מוליך עצבי אינהיבטורי נפוץ מאוד והסיבה של הפחתה זו משוערת להיות חלק מהשפעות ירידת ריכוזי האנזים המקטלז GABA עם הזקנה. בנוסף הנוירונים בקליפה השמיעתית הראשונית (A1) בעכברים מבוגרים הציגו פחות סלקטיביות לתדרים ספציפיים[77], ונצפו גם שינויים בירי הספונטני (Spontaneous firing rate) של התאים[78].
עיבוד שפה בזקנה עריכה

על אף השינויים הרבים המתחוללים במערכת השמיעה בעקבות ההזדקנות, היכולת ביומיום להבין דיבור נשארת יחסית טובה. יכולת זו נשמרת במיוחד כאשר הקלט נקי, כלומר אין רעשי רקע ואין כמה קלטים רלוונטיים הנשמעים בו-זמנית[6]. נמצא כי מבוגרים משתמשים בצורה מעולה בידע שפתי השמור אצלם בזיכרון על מנת להצליח להתגבר על המכשולים שמעמידים הפתלוגיות במערכת השמע. לדוגמה נמצא שימוש בהקשר הסמנטי על מנת להבין מילים שאינן ברורות[80][81], כמו גם שימוש במבנה התחבירי של המשפט על מנת לצפות למילים מסוימות על פני אחרות[82].
ברמה המוחית נמצא שימוש במערכות מוחיות, אשר אינן נמצאות בשימוש הרגיל בעיבוד שפה, על מנת לפצות על היכולות שנפגמות בעיבוד הקלט. לדוגמה הרשת הסינגולו-אופרקולרית (Cingulo-opercular network), רשת המעורבת בתהליכי קשב של עיבוד מלמעלה-למטה ושל שליטה קוגניטיבית, נכנסת לפעולה כאשר מבוגרים מאזינים לדיבור שהוא בעל איכות אקוסטית ירודה[79][83][84]. אזור חשוב נוסף המופיע אצל מבוגרים המשמש לפעילות הפיצוי הוא קליפת המוח הקדם-מוטורית. כאשר מגיע קלט שמיעתי בעל איכות ירודה האזור מופעל ביתר שאת, ולפעילות זו, המערבת אזורים מוטוריים דווקא, ניתנו שני הסברים מרכזיים: האחד הוא הצורך בחזרה על מידע שמיעתי כאשר איכות המידע ירודה[85], השני הוא הצורך בשימוש בתבניות הגיה על מנת להבין דיבור הקשה להבנה[6].
ראו גם עריכה
לקריאה נוספת עריכה
- N.R. Carlson, Physiology of Behavior, 8th ed. (Allyn & Bacon 2004)
קישורים חיצוניים עריכה
 ענר אוטולנגי, רעש פוגע בשמיעה, אבל למה זה קורה?, באתר הארץ, 10 במרץ 2022
ענר אוטולנגי, רעש פוגע בשמיעה, אבל למה זה קורה?, באתר הארץ, 10 במרץ 2022 מסלולים שמיעתיים, דף שער בספרייה הלאומית
מסלולים שמיעתיים, דף שער בספרייה הלאומית
הערות שוליים עריכה
- ^ Schnupp, J., Nelken, I., & King, A. (2011). Auditory neuroscience: Making sense of sound. MIT press.
- ^ 1 2 3 4 5 6 7 8 9 10 11 N.R. Carlson, Physiology of Behavior, 8th ed. (Allyn & Bacon 2004). pages 203-218
- ^ 1 2 3 4 5 6 י. גושן-גוטשטיין, ד. זכאי, נ. גרונאו, פסיכולוגיה קוגניטיבית, האוניברסיטה הפתוחה (2002). כרך ב' עמודים 141.
- ^ 1 2 3 Fettiplace, R., & Hackney, C. M. (2006). The sensory and motor roles of auditory hair cells. Nature reviews neuroscience, 7(1), 19-29.
- ^ Schwander, M., Kachar, B., & Müller, U. (2010). Review series: The cell biology of hearing. The Journal of cell biology, 190(1), 9.
- ^ 1 2 3 4 Peelle, J. E., & Wingfield, A. (2016). The neural consequences of age-related hearing loss. Trends in neurosciences, 39(7), 486-497.
- ^ Ashmore J. 2008. Cochlear outer hair cell mobility. Physiol Rev 88: 173–210.
- ^ Nam JH, Fettiplace R. 2010. Force transmission in the organ of Corti micromachine. Biophys J 98: 2813–2821.
- ^ Kiang, N. Y. S. (1965). Stimulus coding in the auditory nerve and cochlear nucleus. Acta Oto-Laryngologica, 59(2-6), 186-200.
- ^ Pijl, S., & Schwarz, D. W. (1995). Melody recognition and musical interval perception by deaf subjects stimulated with electrical pulse trains through single cochlear implant electrodes. The Journal of the Acoustical Society of America, 98(2), 886-895.
- ^ Pijl, S., & Schwarz, D. W. (1995). Intonation of musical intervals by musical intervals by deaf subjects stimulated with single bipolar cochlear implant electrodes. Hearing Research, 89(1-2), 203-211.
- ^ Brown, C. H., Beecher, M. D., Moody, D. B., & Stebbins, W. C. (1978). Localization of pure tones by Old World monkeys. The Journal of the Acoustical Society of America, 63(5), 1484-1492.
- ^ Perrott, D. R., & Saberi, K. (1990). Minimum audible angle thresholds for sources varying in both elevation and azimuth. The Journal of the Acoustical Society of America, 87(4), 1728-1731.
- ^ Oxenham, A. J., Oxenham, A. J., Plack, C. J., Popper, A. N., & Fay, R. R. (2005). Pitch: neural coding and perception.
- ^ Tomlinson, R. W., & Schwarz, D. W. (1988). Perception of the missing fundamental in nonhuman primates. The Journal of the Acoustical Society of America, 84(2), 560-565.
- ^ Cynx, J., & Shapiro, M. (1986). Perception of missing fundamental by a species of songbird (Sturnus vulgaris). Journal of Comparative Psychology, 100(4), 356.
- ^ Heffner, H., & Whitfield, I. C. (1976). Perception of the missing fundamental by cats. The Journal of the Acoustical Society of America, 59(4), 915-919.
- ^ Webster, D. B., & Fay, R. R. (Eds.). (2013). The mammalian auditory pathway: neuroanatomy (Vol. 1). Springer Science & Business Media.
- ^ Srinivasan, S., Keil, A., Stratis, K., Carr, K. W., & Smith, D. W. (2012). Effects of cross-modal selective attention on the sensory periphery: cochlear sensitivity is altered by selective attention. Neuroscience, 223, 325-332.
- ^ Lakatos, P., Musacchia, G., O’Connel, M. N., Falchier, A. Y., Javitt, D. C., & Schroeder, C. E. (2013). The spectrotemporal filter mechanism of auditory selective attention. Neuron, 77(4), 750-761.
- ^ 1 2 Da Costa, S., van der Zwaag, W., Marques, J. P., Frackowiak, R. S., Clarke, S., & Saenz, M. (2011). Human primary auditory cortex follows the shape of Heschl's gyrus. Journal of Neuroscience, 31(40), 14067-14075.
- ^ 1 2 Bendor, D., & Wang, X. (2006). Cortical representations of pitch in monkeys and humans. Current opinion in neurobiology, 16(4), 391-399.
- ^ Rauschecker, J. P., & Tian, B. (2000). Mechanisms and streams for processing of “what” and “where” in auditory cortex. Proceedings of the National Academy of Sciences, 97(22), 11800-11806.
- ^ 1 2 Terao, Y., Mizuno, T., Shindoh, M., Sakurai, Y., Ugawa, Y., Kobayashi, S., ... & Mochizuki, H. (2006). Vocal amusia in a professional tango singer due to a right superior temporal cortex infarction. Neuropsychologia, 44(3), 479-488.
- ^ Lomber, S. G., & Malhotra, S. (2008). Double dissociation of'what'and'where'processing in auditory cortex. Nature neuroscience, 11(5), 609.
- ^ Poliva, O. (2015). From where to what: A neuroanatomically based evolutionary model of the emergence of speech in humans. F1000Research, 4, 67.
- ^ Klinge, C., Eippert, F., Röder, B., & Büchel, C. (2010). Corticocortical connections mediate primary visual cortex responses to auditory stimulation in the blind. Journal of Neuroscience, 30(38), 12798-12805.
- ^ Schneider, P., Scherg, M., Dosch, H. G., Specht, H. J., Gutschalk, A., & Rupp, A. (2002). Morphology of Heschl's gyrus reflects enhanced activation in the auditory cortex of musicians. Nature neuroscience, 5(7), 688-694.
- ^ Steinbeis, N., Koelsch, S., & Sloboda, J. A. (2006). The role of harmonic expectancy violations in musical emotions: Evidence from subjective, physiological, and neural responses. Journal of cognitive neuroscience, 18(8), 1380-1393.
- ^ Deouell, L. Y. (2007). The frontal generator of the mismatch negativity revisited. Journal of Psychophysiology, 21(3-4), 188-203.
- ^ ראו שיטות ב-Rubin, J., Ulanovsky, N., Nelken, I., & Tishby, N. (2016). The representation of prediction error in auditory cortex. PLoS computational biology, 12(8), e1005058.
- ^ Escera, C., & Corral, M. J. (2007). Role of mismatch negativity and novelty-P3 in involuntary auditory attention. Journal of Psychophysiology, 21(3-4), 251-264.
- ^ Garrido, M. I., Friston, K. J., Kiebel, S. J., Stephan, K. E., Baldeweg, T., & Kilner, J. M. (2008). The functional anatomy of the MMN: a DCM study of the roving paradigm. Neuroimage, 42(2), 936-944.
- ^ Näätänen, R., Paavilainen, P., Rinne, T., & Alho, K. (2007). The mismatch negativity (MMN) in basic research of central auditory processing: a review. Clinical neurophysiology, 118(12), 2544-2590.
- ^ Winkler, I. (2007). Interpreting the mismatch negativity. Journal of Psychophysiology, 21(3-4), 147-163.
- ^ Parras, G. G., Nieto-Diego, J., Carbajal, G. V., Valdés-Baizabal, C., Escera, C., & Malmierca, M. S. (2017). Neurons along the auditory pathway exhibit a hierarchical organization of prediction error. Nature communications, 8(1), 2148.
- ^ Näätänen, R., Paavilainen, P., Titinen, H., Jiang, D., & Alho, K. (1993). Attention and mismatch negativity. Psychophysiology, 30(5), 436-450.
- ^ 1 2 3 4 Bekinschtein, T. A., Dehaene, S., Rohaut, B., Tadel, F., Cohen, L., & Naccache, L. (2009). Neural signature of the conscious processing of auditory regularities. Proceedings of the National Academy of Sciences, 106(5), 1672-1677.
- ^ Brandmeyer, A., Sadakata, M., Spyrou, L., McQueen, J. M., & Desain, P. (2013). Decoding of single-trial auditory mismatch responses for online perceptual monitoring and neurofeedback. Frontiers in neuroscience, 7, 265.
- ^ Dürschmid, S., Reichert, C., Hinrichs, H., Heinze, H. J., Kirsch, H. E., Knight, R. T., & Deouell, L. Y. (2019). Direct evidence for prediction signals in frontal cortex independent of prediction error. Cerebral Cortex, 29(11), 4530-4538.
- ^ 1 2 Salimpoor, V. N., Benovoy, M., Longo, G., Cooperstock, J. R., & Zatorre, R. J. (2009). The rewarding aspects of music listening are related to degree of emotional arousal. PloS one, 4(10), e7487.
- ^ Guhn, M., Hamm, A., & Zentner, M. (2007). Physiological and musico-acoustic correlates of the chill response. Music Perception, 24(5), 473-484.
- ^ Poeppel, D., Emmorey, K., Hickok, G., & Pylkkänen, L. (2012). Towards a new neurobiology of language. Journal of Neuroscience, 32(41), 14125-14131.
- ^ Peelle, J. E., & Davis, M. H. (2012). Neural oscillations carry speech rhythm through to comprehension. Frontiers in psychology, 3, 320.
- ^ Rimmele, J. M., Golumbic, E. Z., Schröger, E., & Poeppel, D. (2015). The effects of selective attention and speech acoustics on neural speech-tracking in a multi-talker scene. Cortex, 68, 144-154.
- ^ Golumbic, E. M. Z., Ding, N., Bickel, S., Lakatos, P., Schevon, C. A., McKhann, G. M., ... & Poeppel, D. (2013). Mechanisms underlying selective neuronal tracking of attended speech at a “cocktail party”. Neuron, 77(5), 980-991.
- ^ Davis, M. H., & Johnsrude, I. S. (2007). Hearing speech sounds: top-down influences on the interface between audition and speech perception. Hearing research, 229(1-2), 132-147.
- ^ Maqbool, M., & Maqbool, S. (2013). Textbook of Ear, Nose and Throat Diseases. JP Medical Ltd.
- ^ Kumar, S., Sedley, W., Barnes, G. R., Teki, S., Friston, K. J., & Griffiths, T. D. (2014). A brain basis for musical hallucinations. Cortex, 52, 86-97.
- ^ Lanting, C. P., De Kleine, E., & Van Dijk, P. (2009). Neural activity underlying tinnitus generation: results from PET and fMRI. Hearing research, 255(1-2), 1-13.
- ^ Ropper, A. H., Adams, R., Victor, M., & Samuels, M. A. (2005). Adams and Victor's principles of neurology. McGraw Hill Medical,. Chapter 1
- ^ Anderson, S., Parbery-Clark, A., White-Schwoch, T., & Kraus, N. (2012). Aging affects neural precision of speech encoding. Journal of Neuroscience, 32(41), 14156-14164.
- ^ Cruickshanks, K. J., Wiley, T. L., Tweed, T. S., Klein, B. E., Klein, R., Mares-Perlman, J. A., & Nondahl, D. M. (1998). Prevalence of hearing loss in older adults in Beaver Dam, Wisconsin: The epidemiology of hearing loss study. American journal of epidemiology, 148(9), 879-886.
- ^ Plack, C. J., Barker, D., & Prendergast, G. (2014). Perceptual consequences of “hidden” hearing loss. Trends in hearing, 18, 2331216514550621.
- ^ Fellinger, J., Holzinger, D., Gerich, J., & Goldberg, D. (2007). Mental distress and quality of life in the hard of hearing. Acta Psychiatrica Scandinavica, 115(3), 243-245.
- ^ Gates, G. A., & Mills, J. H. (2005). Presbycusis. The lancet, 366(9491), 1111-1120.
- ^ Humes, L. E., Kewley-Port, D., Fogerty, D., & Kinney, D. (2010). Measures of hearing threshold and temporal processing across the adult lifespan. Hearing research, 264(1-2), 30-40.
- ^ Kujawa, S. G., & Liberman, M. C. (2009). Adding insult to injury: cochlear nerve degeneration after “temporary” noise-induced hearing loss. Journal of Neuroscience, 29(45), 14077-14085.
- ^ Sergeyenko, Y., Lall, K., Liberman, M. C., & Kujawa, S. G. (2013). Age-related cochlear synaptopathy: an early-onset contributor to auditory functional decline. Journal of Neuroscience, 33(34), 13686-13694.
- ^ Kujawa, S. G., & Liberman, M. C. (2015). Synaptopathy in the noise-exposed and aging cochlea: Primary neural degeneration in acquired sensorineural hearing loss. Hearing research, 330, 191-199.
- ^ Bao, J., & Ohlemiller, K. K. (2010). Age-related loss of spiral ganglion neurons. Hearing research, 264(1-2), 93-97.
- ^ Gray, D. T., Engle, J. R., & Recanzone, G. H. (2014). Age‐related neurochemical changes in the rhesus macaque cochlear nucleus. Journal of Comparative Neurology, 522(7), 1527-1541.
- ^ Engle, J. R., Gray, D. T., Turner, H., Udell, J. B., & Recanzone, G. H. (2014). Age-related neurochemical changes in the rhesus macaque inferior colliculus. Frontiers in aging neuroscience, 6, 73.
- ^ Caspary, D. M., Ling, L., Turner, J. G., & Hughes, L. F. (2008). Inhibitory neurotransmission, plasticity and aging in the mammalian central auditory system. Journal of Experimental Biology, 211(11), 1781-1791.
- ^ Walton, J. P., Simon, H., & Frisina, R. D. (2002). Age-related alterations in the neural coding of envelope periodicities. Journal of neurophysiology, 88(2), 565-578.
- ^ Strouse, A., Ashmead, D. H., Ohde, R. N., & Grantham, D. W. (1998). Temporal processing in the aging auditory system. The Journal of the Acoustical Society of America, 104(4), 2385-2399.
- ^ Schneider, B. A., Pichora‐Fuller, M. K., Kowalchuk, D., & Lamb, M. (1994). Gap detection and the precedence effect in young and old adults. The Journal of the Acoustical Society of America, 95(2), 980-991.
- ^ Walton, J. P. (2010). Timing is everything: temporal processing deficits in the aged auditory brainstem. Hearing research, 264(1-2), 63-69.
- ^ Pichora-Fuller, M. K., & Souza, P. E. (2003). Effects of aging on auditory processing of speech. International journal of audiology.
- ^ Gordon-Salant, S., Yeni-Komshian, G. H., Fitzgibbons, P. J., & Barrett, J. (2006). Age-related differences in identification and discrimination of temporal cues in speech segments. The Journal of the Acoustical Society of America, 119(4), 2455-2466.
- ^ Skoe, E., Krizman, J., Anderson, S., & Kraus, N. (2015). Stability and plasticity of auditory brainstem function across the lifespan. Cerebral Cortex, 25(6), 1415-1426.
- ^ Ng, C. W., Navarro, X., Engle, J. R., & Recanzone, G. H. (2015). Age-related changes of auditory brainstem responses in nonhuman primates. Journal of neurophysiology, 114(1), 455-467.
- ^ Burianova, J., Ouda, L., Profant, O., & Syka, J. (2009). Age-related changes in GAD levels in the central auditory system of the rat. Experimental gerontology, 44(3), 161-169.
- ^ Ling, L. L., Hughes, L. F., & Caspary, D. M. (2005). Age-related loss of the GABA synthetic enzyme glutamic acid decarboxylase in rat primary auditory cortex. Neuroscience, 132(4), 1103-1113.
- ^ Profant, O., Balogová, Z., Dezortová, M., Wagnerová, D., Hájek, M., & Syka, J. (2013). Metabolic changes in the auditory cortex in presbycusis demonstrated by MR spectroscopy. Experimental gerontology, 48(8), 795-800.
- ^ Gao, F., Wang, G., Ma, W., Ren, F., Li, M., Dong, Y., ... & Edden, R. A. (2015). Decreased auditory GABA+ concentrations in presbycusis demonstrated by edited magnetic resonance spectroscopy. Neuroimage, 106, 311-316.
- ^ Turner, J. G., Hughes, L. F., & Caspary, D. M. (2005). Affects of aging on receptive fields in rat primary auditory cortex layer V neurons. Journal of neurophysiology, 94(4), 2738-2747.
- ^ Hughes, L. F., Turner, J. G., Parrish, J. L., & Caspary, D. M. (2010). Processing of broadband stimuli across A1 layers in young and aged rats. Hearing research, 264(1-2), 79-85.
- ^ 1 2 Vaden, K. I., Kuchinsky, S. E., Ahlstrom, J. B., Dubno, J. R., & Eckert, M. A. (2015). Cortical activity predicts which older adults recognize speech in noise and when. Journal of Neuroscience, 35(9), 3929-3937.
- ^ Benichov, J., Cox, L. C., Tun, P. A., & Wingfield, A. (2012). Word recognition within a linguistic context: Effects of age, hearing acuity, verbal ability and cognitive function. Ear and Hearing, 32(2), 250.
- ^ Lash, A., Rogers, C. S., Zoller, A., & Wingfield, A. (2013). Expectation and entropy in spoken word recognition: Effects of age and hearing acuity. Experimental Aging Research, 39(3), 235-253.
- ^ DeCaro, R., Peelle, J. E., Grossman, M., & Wingfield, A. (2016). The two sides of sensory–cognitive interactions: Effects of age, hearing acuity, and working memory span on sentence comprehension. Frontiers in psychology, 7, 236.
- ^ Wild, C. J., Yusuf, A., Wilson, D. E., Peelle, J. E., Davis, M. H., & Johnsrude, I. S. (2012). Effortful listening: the processing of degraded speech depends critically on attention. Journal of Neuroscience, 32(40), 14010-14021.
- ^ Vaden, K. I., Kuchinsky, S. E., Cute, S. L., Ahlstrom, J. B., Dubno, J. R., & Eckert, M. A. (2013). The cingulo-opercular network provides word-recognition benefit. Journal of Neuroscience, 33(48), 18979-18986.
- ^ Rönnberg, J., Lunner, T., Zekveld, A., Sörqvist, P., Danielsson, H., Lyxell, B., ... & Rudner, M. (2013). The Ease of Language Understanding (ELU) model: theoretical, empirical, and clinical advances. Frontiers in systems neuroscience, 7, 31.
}}