פְלוּאוֹרֶסְצֶנְצְיָה (או פלואורסצנטיות; באנגלית: Fluorescence; בעברית גם: פְלוּאוֹרָנוּת, זְהִירָה) היא פליטה של אור מחומר שנחשף לאור, לרוב בצבע שונה.
בשונה מסוגים אחרים של לומינסנציה כגון פוספורסצנציה, זמני החיים של תופעה זו קצרים יחסית והם מאופיינים בסדר גודל של ננו-שניות.
בדומה לפיזור ראמאן, גם פלואורסצנציה היא דוגמה להיסט סטוקס אף כי שני התהליכים מביאים לתוצאה זהה — פליטת אור באורך גל גדול יותר ("אדום" יותר) — ישנם 2 הבדלים בין התופעות:
בפלואורסצנציה אין פיזור ואנרגיית העירור נבלעת לחלוטין.
פיזור ראמאן, בניגוד לפלואורסצנציה, אינו מוגבל לטווח תדרים/אורכי גל בבליעה ולכן אינו נחשב תופעה תהודתית.
כוס עץ העשויה מעץ הנארה מלאה בתמצית מעץ הכליות, ובקבוק המכיל את תמיסתו הפלואורסצנטית.
עדות ראשונה לתופעת הפלואורסצנציה תוארה ב-1560 על ידי ברנרדינו דה סאגון בקודקס הפלורנטיני וב-1565 על ידי ניקולס מונדרס בהקשר לתרופה ששימשה כחומר משתן והופקה מעץ שכונה "עץ הכליות".[1][2][3][4] התרופה נודעה בתכונתה לשנות את הצבע של המים בהתאם לאור ולזווית פגיעתו.
אני כמעט נוטה לצקת מטבע לשון ולקרוא לתופעה זו פלואורסצנציה על שם הפלואוריט, באנלוגיה לתופעת האופאלסצנציה ששמה נגזר משם המינרל, אופאל
המקור באנגלית
I am almost inclined to coin a word, and call the appearance fluorescence, from fluor-spar [i.e., fluorite], as the analogous term opalescence is derived from the name of a mineral.
דיאגרמת יבלונסקי של מולקולה אורגנית. ציר ה-Y מבטא אנרגיה ביחס למצב היסוד של המולקולה, קווים מלאים מתארים מעברים קרינתיים (מלווים בליעה או פליטה של פוטון) וקווים שבורים מתארים מעברים לא קרינתיים. משמאל: מעברים בבליעה מרמת היסוד לרמות סינגלט מעוררות (כיוון שמהמעבר האלקטרוני מהיר מסידור האטומים המולקולה תהיה במצב מעורר ויברציונית מיד לאחר הערור), במרכז: מעבר מהמצב המעורר S1 לרמת היסוד (שוב, לרמות ויברציה מעוררות) המלווה בפליטת פוטון - פלואורסצנציה, מימין: דעיכה מרמת טריפלט לרמת היסוד: פוספורסצנציה
מולקולה ב- יכולה לעבור רלקסציה בכמה תהליכים שונים מתחרים:
המולקולה יכולה לעבור תהליך לא-קרינתי שבו אנרגיית העירור הופכת לחום (ויברציות) על הממס.
מולקולות אורגניות מעוררות יכולות גם לעבור דעיכה לא קרינתית של אלקטרון ממצב סינגלט מעורר למצב טריפלט בתהליך שנקרא חצייה בין-מערכתית (Intersystem crossing). במקרה זה, הרלקסציה נעשית או באמצעות פליטתפוטון בתהליך שנקרא פוספורסנציה (זַרְחוֹרָנוּת, פוֹסְפוֹרֶסֶנְטִיּוּת) או על ידי הפיכה לחום בתהליך לא -קרינתי משני.
ברוב המקרים, לאור הנפלט יש אורך גל ארוך יותר, ולכן אנרגיה נמוכה יותר, מאשר הקרינה שנספגה. תופעה זו ידועה בשם היסט סטוקס. עם זאת, כאשר הקרינה האלקטרומגנטית הנספגת חזקה, מתאפשר לאלקטרון אחד לבלוע שני פוטונים. בליעת דו פוטונית (אנ') זו יכולה להוביל לפליטת קרינה בעלת אורך גל קצר יותר מאשר הקרינה הנספגת. הקרינה הנפלטת עשויה גם להיות באותו אורך גל כמו קרינה נספגת, המכונה "קרינת תהודה".[10]
מולקולותמעוררות באמצעות בליעת אור או תהליך אחר (למשל כתוצר של תגובה) יכולות להעביר אנרגיה למולקולה "מעוררת" שנייה, אשר מומרת למצב המעורר שלה ויכולה לפלוט אור פלואורסצנטי.
דיאגרמת יבלונסקי מתארת את רוב מנגנוני הרלקסציה למולקולות במצב מעורר. התרשים מראה כיצד פלואורסצנציה מתרחשת עקב רלקסציה של אלקטרונים מעוררים מסוימים של המולקולה.[11]
הנצילות הקוונטית הפלואורסצנטית מבטאת את היעילות של תהליך פלואורסצנטי. הנצילות מוגדרת כיחס בין מספר הפוטונים שנפלטו לבין מספר הפוטונים שנבלעו:[12][13]
הנצילות הקוונטית הפלואורסצנטית המקסימלית האפשרית היא 1 (100%) - במצב זה, כנגד כל פוטון שנבלע יש פוטון שנפלט. גם תרכובות עם נצילות קוונטית נמוכה של 0.1 (10%) עדיין נחשבות פלואורסצנטיות.
דרך נוספת להגדיר את הנצילות הקוונטית של הקרינה היא על ידי קצב הדעיכה של המולקולה מהרמה המעוררת לרמת היסוד:
כאשר הוא ריכוז המולקולות במצב מעורר בזמן , הוא ריכוז המולקולות במצב מעורר ההתחלתי (בזמן ) ו-הוא קצב הדעיכה ההופכי לזמן החיים הפלואורסצנטי, .
זוהי דוגמה קלאסית לדעיכה מעריכית. תהליכים קרינתיים ולא קרינתיים שונים יכולים לגרום לאי אכלוס של הרמה המעוררת ולכן קצב הדעיכה הכללי, , הוא הסכום של קצב התהליכים הקרינתיים, וקצב התהליכי הלא קרינתיים :
עבור תרכובות פלואורסצנטיות נפוצות, טווח זמני הרלקסציה האופייניים עבור פליטת פוטון מ-UV עד אינפרא אדום, נע בין 0.5 עד 20 ננו-שניות. זמן חיים פלואורסצנטי הוא פרמטר חשוב עבור יישומים מעשיים כגון: FRET ו-FLIM (אנ').
פלואורופורים נוטים יותר להיות מעוררים על ידי פוטונים אם מומנט המעבר הדיפולי של הפלואורופור מקביל לווקטור החשמלי של הפוטון.[12] גם הקיטוב של האור הנפלט תלוי במומנט המעבר. מומנט המעבר הדיפולי תלוי בכיוון הפיזי של מולקולת הפלואורופור. עבור מולקולה הנתונה בתמיסה זה אומר כי עוצמת הקיטוב של האור הנפלט תלויה בדיפוזיה סיבובית. לכן, ניתן להשתמש במדידות אנאיזוטרופיות כדי לחקור תנועה של מולקולה בסביבה מסוימת.
כמותית ניתן להגדיר אנאיזוטרופיות כך:
כאשר היא העוצמה הנפלטת בכיוון המקביל לקיטוב האור המעורר ואילו היא העוצמה נפלטת בכיוון מאונך לקיטוב האור המעורר.[14]
סכמה של חוק קאשה. פוטון עם אנרגיה מעורר אלקטרון של רמת הבסיס, של אנרגיה , עד לרמת אנרגיה מעוררת (או ) או עד לאחת מרמות המשנה הויברציוניות. תרשים לאנרגיית פרנק קונדון. בורות הפוטנציאל מראים שהמעבר העדיף הוא בין v = 0 לבין v = 2.מעברי הרמות בפיזור ראמאן.
החוק קובע כי הנצילות הקוונטית של לומינסציה אינה תלויה באורך הגל של קרינת העירור.[15] הסיבה לתופעה נעוצה בכך שבמולקולה מעוררת בדרך כלל מתרחשת דעיכה לרמה הוירבציונית הנמוכה ביותר של רמת העירור בטרם מתרחשת פלואורסצנציה. חוק זה ניתן לניסוח באופן אחר: ספקטרום הפלואורסצנציה כמעט ואינו תלוי באורך הגל של קרינת העירור.
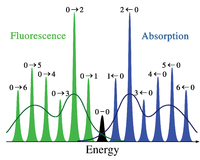
ערך מורחב – היסט סטוקס ייצוג סכמטי של ספקטרום הבליעה והפלואורסצנציה על פי אנרגיית פנרק קונדון הסימטריה נובעת מהצורה השווה של בורות הפוטנציאל ברמת היסוד וברמה המעוררת.
באופן כללי, לאור הנפלט בפלואורסצנציה יש אורך גל ארוך יותר ואנרגיה נמוכה יותר מהאור הנבלע. תופעה זו, הידועה בשם היסט סטוקס, נובעת מאובדן אנרגיה בין הזמן שבו הפוטון נבלע, לבין הזמן שבו הוא נפלט. הגורמים וסדר הגודל של היסט סטוקס יכולים להיות מורכבים והם תלויים בפלואורופור ובסביבתו. עם זאת, יש כמה סיבות נפוצות. לעיתים קרובות ישנה דעיכה לא קרינתית לרמה הויברציונית הנמוכה ביותר של רמת העירור. גורם נוסף הוא שהפליטה מותירה את הפלואורופור ברמה ויברציונית גבוהה יותר מרמת היסוד.
התופעה מוגדרת כבליעת גלים אלקטרומגנטיים בתחום האור הנראה על ידי חלבונים פלואורסצנטיים באורגניזם חי ופליטת אור באנרגיה נמוכה יותר. צבע האור הנפלט שונה מצבע האור הנבלע. האלקטרון המעורר, עולה לרמה לא יציבה. חוסר היציבות של הרמה גבוה מאוד שכן חזרת האלקטרון לרמת היסוד היציבה מתרחשת כמעט מיידית. . חזרה זו תואמת לשחרורו של עודף אנרגיה בצורה של אור פלואורסצנטי. פליטת אור זו ניתנת לצפייה רק כאשר האור הממריץ עדיין מספק אור לאורגניזם והוא בדרך כלל צהוב, ורוד, כתום, אדום, ירוק או סגול.
ביופלואורסצנציה בתגובה לאור אולטרה סגול נמצאה במיני יונקים רבים, רובם פעילי לילה או פעילי דמדומים. אך יש גם פעילי יום זוהרים, כגון זברה ודוב קוטב. לכל היונקים - כולל האדם - שיניים זוהרות[16].
תופעה בה אנרגיה ממקור אור חיצוני נקלטת על ידי חלבונים פלואורסצנטיים באורגניזם חי ומשוחררת זמן מה לאחר מכן באורך גל ארוך יותר. בניגוד לביופלואורסצנציה זמן החיים של התופעה ארוך יותר.
תאיפיגמנט המציגים פלואורסצנציה נקראים כרומטופורים פלורוסצנטיים, והם מתפקדים באופן דומה לכרומטופורים רגילים. תאים אלה הם דנדריטים, ומכילים פיגמנטים הנקראים פלואורוזומים. פיגמנטים אלה מכילים חלבונים פלואורסצנטיים המופעלים על ידי יוני K + (אשלגן). התנועה, הצבירה והפיזור שלהם בתוך הכרומטופור הזוהר הם הגורמים לדפוס פלואורסצנטי מכוון.[17][18] כמו כרומטופורים אחרים כגון מלנופורים, גם תאי פיגמנט מכילים מלנין. מערכת העצבים נשלטת על ידי דפוס פלואורסצנטי לטווח קצר.[17] ניתן למצוא כרומטופורים פלואורסצנצטיים בעור (למשל בדגים) ממש מתחת לאפידרמיס.
מיםבולעיםאור באורכי גל ארוכים כך שפחות אור מאורכי הגל הללו משתקף חזרה ומגיע לעין. לכן, צבעים חמים מספקטרום האור הנראה נראים פחות תוססים בעומקים הולכים וגוברים. מים מפזרים אור באורך גל קצר יותר מעל סגול, כלומר צבעים קרירים יותר חולשים על שדה הראייה באזור הצילום. עוצמת האור פוחתת פי 10 עם כל 75 מטר של עומק, כך שבעומק של 75 מטר, עוצמת האור היא % 10 מהעוצמה על פני השטח, ואילו בעומק של 150 מטר העוצמה היא רק % 1 מהעוצמה על פני השטח. בגלל סינון זה של האור, ניתן לראות חומרים שונים בעומקים שונים.[19][20][21]
לדגי גרם החיים במים רדודים בדרך כלל יש ראיית צבע טובה בגלל מגוריהם בסביבה צבעונית. לפיכך, בדגי מים רדודים, פלואורסצנציה אדומה, כתומה וירוקה משמשת ככל הנראה אמצעי לתקשורת.[19]
דגים רבים המציגים פלואורסצנציה, כמו כרישים, דגי לטאה, דגי עקרב, דגי נוי ודגים שטוחים, הם בעלי מסנן תוך-עיני צהוב. מסננים אלו מאפשרים לדגים לשפר את הניגודיות והדפוסים הוויזואליים שלא נראים לעין דגים וטורפים אחרים חסרי תכונה זו.[22]
פלואורסצנציה משמשת למגוון רחב של פונקציות באלמוגים. חלבונים פלואורסצנטיים באלמוגים עשויים לתרום לפוטוסינתזה על ידי המרת אורכי גל שאינם שמישים אחרת לאלה שעבורם האצות הסימביוטיות של האלמוגים מסוגלות לבצע פוטוסינתזה.[23] כמו כן, החלבונים עשויים להשתנות במספרם ע"פ מידת האור הנבלע. [39] באופן דומה, חלבונים פלואורסצנטיים אלו עשויים להיות בעלי יכולות נוגדות חמצון כדי לחסל רדיקליחמצן המיוצרים על ידי פוטוסינתזה.[24] במילים אחרות, באמצעות אפנוןפוטוסינתזה, החלבונים הפלואורסצנטיים משמשים כאמצעי לוויסות הפעילות של סימביוני האצות הפוטוסינתטיים של האלמוגים.[25]
Aequorea victoria, מדוזה ביו-פלואורסצנטית הידועה כנשאית GFP
ל- Subulata Alloteuthis ו- Loligo vulgaris, שני סוגים של דיונונים כמעט שקופים, יש כתמים פלואורסצנטיים מעל עיניהם. כתמים אלה מחזירים את האור הפוגע ומשמשים לצורכי הסוואה ותקשורת.[26]
דוגמה נוספת ידועה לפלואורסצנציה באוקיינוס היא ההידרה Aequorea victoria. מדוזה זו חיה באזור מול חופה המערבי של צפון אמריקה וזוהתה כנשא של חלבון פלואורסצנטי ירוק (GFP) על ידי אוסאמו שיממורה. הגן של חלבונים פלואורסצנטיים ירוקים משמעותי מבחינה מדעית במחקרים גנטיים כדי להצביע על ביטוים של גנים אחרים.[27]
לכמה מינים של טרפידים יש סימנים פלואורסצנטיים צהובים לאורך קשקשתם לצורכי הגנה מפני טורפים. במהלך טקסי הזדווגות, הטרפיד זוהר באופן פעיל ופולט אורך גל שיכול להקלט אצל טרפידים אחרים.[28]
צפרדע העץ נקודת פולקה (אנ'), הנפוצה באמזונאס, התגלתה כדו חיים הפלואורסצנטי הראשון בשנת 2017. עור הצפרדע הוא בעל צבע ירוק בהיר עם נקודות בצבע לבן, צהוב או אדום בהיר. הפלואורסצנציה של הצפרדע התגלתה שלא במתכוון בבואנוס איירס, ארגנטינה. את הפלואורסצנצי ניתן לייחס לתרכובת חדשה שנמצאה בלימפה ובעור.[30] התרכובת הפלואורסצנטית העיקרית היא Hyloin-L1 והיא מעניקה זוהר כחול-ירוק כאשר היא נחשפת לאור סגול או על-סגול. מדענים העומדים מאחורי התגלית מציעים כי הפלואורסצנציה משמשת לצורכי תקשורת. הם גם מעריכים כי כמאה או 200 מינים של צפרדעים עשויים להיות פלורסצנטיים.[31]
לתוכאים יש פלומת פלואורסצנטית שעשויה לשמש באיתות בתהליך זיווג. מחקר שנערך בתוכונים מצויים הראה כי זכרים ונקבות העדיפו משמעותית ציפורים עם הפיגמנט הפלואורסצנטי. ממחקר זה עולה כי הפלומה הקורנת של התוכיים אינה סתם תוצר לוואי של פיגמנטציה, אלא איתות מיני מובנה.[33]
פרח הלילנית הרב-גונית (אנ') מכיל פיגמנטים צהובים וסגולים פלואורסצנטיים. תחת אור לבן, חלקים מהפרח המכילים רק בטא-קסנטינים נראים צהובים, אך באזורים בהם קיימים גם בטא-קסנטינים וגם בטא-ציאנינים, הפלואורסצנציה דוהה בגלל מנגנוני סינון פנימיים.[36]
הכלורופיל הוא ככל הנראה המולקולה הפלואורסצנטית הנפוצה ביותר ומייצרת פליטה אדומה תחת מנעד רחב של אורכי גל מעוררים.[37]
הפלואורסצנציה במינרלים נגרמת על ידי מגוון רחב של אקטיבטורים (אנ'). במקרים מסוימים, יש להגביל את ריכוז האקטיבטור מתחת לרמה מסוימת, כדי למנוע רוויה ודיכוי של פליטת הפלואורסצנציה. יתר על כן, המינרל צריך להיות נקי מזיהומים כמו ברזל או נחושת, כדי למנוע דיכוי פלואורסצנטי. מנגן דיולנטי, בריכוזים של עד אחוזים בודדים, אחראי לפלואורסצנציה האדומה או הכתומה של קלציט, לפלואורסצנציה הירוקה של וילמיט, לפלואורסצנציה הצהובה של האספריט ולפלואורסצנציה הכתומה של וולסטוניט וקלינוהדריט. אורניום הקסולנטי, זוהר בכל הריכוזים בצבע ירוק צהוב, והוא הגורם לפלואורסצנציהשל מינרלים כמו אוטוניט או אנדרסונייט. כרום טריולינטי בריכוז נמוך הוא המקור לפלואורסצנציה האדומה של אודם. אירופיום דיולנטי הוא המקור לפלואורסצנציה הכחולה, כאשר הוא נראה במינרל הפלואוריט. לנתנידים טריולינטיים כמו טרביום ודיספרוסריום הם הגורמים העיקריים לפלואורסצנציה הצהובה שמפגין המינרל פלואור, ותורמים לפלואורסצנציה הכתומה של זירקון. סידן מוליבדאט וסידן טונגסטייט זוהרים באופן מובהק בצהוב וכחול, בהתאמה. כאשר הם נמצאים יחד בתמיסה מוצקה, מועברת אנרגיה מהטונגסטן בעלת האנרגיה הגבוהה יותר למוליבדן באנרגיה נמוכה יותר, כך שרמות נמוכות למדי של מוליבדן מספקות כדי לגרום לפליטה צהובה עבור סידן טונגסטייט, במקום כחול. ספלריט (אבץ גופרתי), זוהר במגוון צבעים, המושפעים מנוכחות זיהומים שונים.
נפט גולמי זוהר במגוון צבעים, החל מחום עמום לשמנים כבדים ועד לצבעי צהבהב בהיר וכחלחל-לבן לשמנים קלים. תופעה זו משמשת בקידוחים וחיפושי נפט לזיהוי כמויות קטנות מאוד של שמן.
תמיסות אורגניות כגון אנתרסן או סטילבן, מומסים בבנזן או טולואן, זוהרים עם הקרנת אולטרה סגול או קרני גמא. זמני ההתפרקות של פלואורסצנציה זו הם בסדר גודל של ננו-שניות, מכיוון שמשך ההארה תלוי בזמן החיים של המצבים המעוררים של החומר הפלואורסצנטי, במקרה זה אנתרסן או סטילבן.[38]
ציור ופלסטיק מוארים על ידי צינורות UV.פירוק של מקלות זוהרים (אנ')
הנורה הפלואורסצנטית הנפוצה מבוססת על פלואורסצנציה. שפופרת או צינור הזכוכית מכיל ואקום חלקי וכמות קטנה של כספית. פריקה חשמלית בצינור גורמת לאטומי הכספית לפלוט בעיקר אור אולטרה סגול. הצינור מרופד בציפוי של חומר פלואורסצנטי, הנקרא זרחן, אשר סופג את האור האולטרה סגול ופולט מחדש אור בתחום הנראה.
התאורה הפלואורסצנטית יותר חסכונית באנרגיה בהשוואה לתאורה מבוססת אלמנטי להט. עם זאת, ספקטרום לא אחיד של תאורה פלואורסצנטית עלול לגרום לצבעים מסוימים להיראות שונה לעומת תאורת להט או תאורת יום באותו צבע. ספקטרום פליטת אדי הכספית מכיל קו אולטרה סגול קצר גל ב-254 ננומטר (המספק את רוב האנרגיה לזרחנות) וקווי אור נראה ב-436 ננומטר (כחול), 546 ננומטר (ירוק) ו-579 ננומטר (כתום-צהוב). ניתן לראות 3 קווים אלו על גבי הרצף הלבן באמצעות ספקטרוסקופ היד, עבור האור הנפלט על ידי צינורות ניאון לבן רגיל. אותם קווים נראים לעין, בתוספת פליטה של אירופיום וטרביום מהווים את ספקטרום הפליטה שעליו מבוסס עקרון הפעולה של נורה פלואורסצנטית קומפקטית.[40]
תהליכים אנליטיים רבים כוללים שימוש בפלואורומטר, לרוב באורך גלמעורר יחיד ואורך גל לגילוי יחיד. בגלל הרגישות שהשיטה מעניקה, ניתן למדוד ריכוזי מולקולות פלואורסצנטיות בדיוק של עד אחד חלקי טריליון. [42]
מדידת פלואורסצנציה בכמה אורכי גל נעשית על ידי מערך גלאים, כדי לאתר תרכובות מזרימת HPLC. כמו כן, ניתן לבצע ויזואליזציה של פלטות TLC (אנ') אם התרכובת או מגיב הצביעה הוא פלואורופור. הפלואורסצנציה אפקטיבית יותר כאשר יש יחס גדול יותר של אטומים ברמות אנרגיה נמוכות בהתפלגות בולצמן. אם כן, קיימת סבירות גבוהה יותר לעירור ושחרור פוטונים על ידי אטומים בעלי אנרגיה נמוכה יותר, מה שמייעל את התהליך.
בדרך כלל ההתקן של תבחין (אנ') ביולוגי פלואורסצנטי מכיל מקור אור שפולט אורכי גל שונים של אור. באופן כללי, נדרש רק אורך גל יחיד כדי לבצע אנליזה ראויה, ולכן כדי לסנן סלקטיבית את האור הוא עובר דרך מונוכרומטורעירור. בהמשך עובר אורך הגל המסוים דרך תא הדוגמה. אחרי בליעה ורה-פליטה, אורכי גל רבים מתמזגים עקב היסט סטוקס ומעברי אלקטרון. כדי להפריד ולנתח אותם, הקרינה הפלואורסצנטית מועברת דרך מונוכרומטור פליטה ונמדדת סלקטיבית בגלאי.
בתחום חקר המים, שימוש בספקטרוסקופיה פלואוסנטית משמש לזיהוי וניטור של מזהמים אורגניים במים.[43] בזכות פריצות דרך בתחום מדעי המחשב ולימוד מכונה, נמצא שניתן להשתמש בשיטה גם לזיהוי זיהום חיידקי של מי שתייה.[44]
פלואורסצנציה במדעי החיים משמשת ככלל כדרך לא מזיקה למעקב ולניתוח מולקולות ביולוגיות באמצעות פליטה פלואורסצנטית בתדירות ספציפית שבה אין קרינת רקע מהאור המעורר למעט רכיבים בעלי פלואורסצנציה חלשה (פלואורסצנציה עצמית). למעשה, ניתן "לתייג" חלבון או מרכיב אחר עם פלואורופור חיצוני, צבען פלואורסצנטי, שיכול להיות מולקולה קטנה, חלבון או נקודה קוונטית. לתיוג מסוג זה יש שימוש בהרבה יישומים ביולוגיים.[45]
הכימות של צבען נעשה באמצעות ספקטרופלואורמטר ומשמש ביישומים נוספים כגון:
תאים אנדותליים תחת מיקרוסקופ פלואורסצנטי עם 3 ערוצים נפרדים המסמנים רכיבי תא ספציפיים.בעת סריקת עוצמת הפלואורסצנציה על פני משטח נוצרת מיקרוסקופיה פלואורסצנטית של רקמות, תאים או מבנים תת-תאיים, אשר מושגת על ידי תיוג נוגדן עם פלואורופור ומאפשרת לנוגדן למצוא את אנטיגן המטרה שלו בתוך הדגימה. תיוג נוגדנים מרובים עם פלואורופורים שונים מאפשר הדמיה של יעדים מרובים בתמונה יחידה (מספר ערוצים). מיקרו מערכי DNA הם אחת הווריאציות לכך.
אימונולוגיה: נוגדן מוכן מראש על ידי חיבור לקבוצה כימית פלואורסצנטית, עקיבה ואפילו כימות של האתרים אליהם נקשר הנוגדן נעשית על ידי מיקרוסקופ פלואורסצנטי.
(FLIM (Fluorescence-lifetime imaging microscopy - מיקרוסקופית דימות זמן חיים פלואורסצנטי (אנ') - משמשת באיתור אינטראקציות ביו-מולקולריות מסוימות המתבטאות על ידי השפעה על זמן החיים הפלואורסצנטי.
דיאגרמת פעולה של מכשיר ה-FACS (מיון תאים פלואורסצנטי) באמצעות ציטומטרית זרימה (flow cytometry).
FRET (מעבר אנרגיה בתהודה פלואורסצנטית) - משמש כסמן ביולוגי לחקר אינטראקציות חלבון וזיהוי רצפי חומצות גרעין מסוימים. שילוב עם (FLIM (Fluorescence-lifetime imaging microscopy מוסיף עוד שכבת מידע.
ריצוף אוטומטי של DNA בשיטת סיום שרשרת: לכל אחד מארבעה בסיסי סיום שרשרת שונים יש תיוג פלורסצנטי ספציפי משלו. כאשר מופרדים מולקולות ה- DNA המסומנות, התווית הפלואורסצנטית מעוררת על ידי מקור UV, וזהות הבסיס המסיים את המולקולה מזוהה על ידי אורך הגל של האור הנפלט.
fluorescence-activated cell sorting) FACS) - אחת מכמה טכניקות מיון תאים חשובות המשמשות להפרדה של קווי תאים שונים (במיוחד כאלה שמבודדים מרקמות של בעלי חיים).
גילוי DNA: לתרכובת אתידיום ברומיד, בתמיסה מימית, יש מעט מאוד פלואורסצנציה, מכיוון שהיא מרווה על ידי מים. הפלואורסצנציה של האתידיום ברומיד מוגברת מאוד לאחר שהיא נקשרת ל- DNA, ולכן תרכובת זו שימושית מאוד בהמחשת המיקום של שברי DNA באלקטרופורזה של ג'לאגרוז. האתידיום נמצא בסביבה הידרופובית כאשר הוא נמצא בין זוגות הבסיס של ה-DNA, ולכן מוגן מפני ליבון פלואורסצנטי על ידי מים. אתידיום ברומיד עשוי להיות מסרטן - אלטרנטיבה בטוחה יותר היא SYBRירוק.
צבעים פלורסצנטיים משמשים לעיתים קרובות בשילוט, במיוחד שלטי דרכים. בדרך כלל ניתן לזהות צבעים פלורסצנטיים בטווחים ארוכים יותר מאשר הצבעים הלא פלואורסצנטיים, כאשר הכתום הפלואורסצנטי בולט במיוחד והוא נמצא בשימוש תכוף בשלטים ותוויות בטיחות.[47]
תרכובות פלואורסצנטיות משמשות לרוב כדי לשפר את מראה הבד והנייר, וגורמות לאפקט "הלבנה". משטח לבן המטופל במבהיר אופטי יכול לפלוט אור זוהר יותר מזה שמאיר עליו, ולגרום לו להיראות בהיר יותר. האור הכחול שנפלט על ידי המבהיר מפצה את הכחול שהולך ומצטמצם של החומר המטופל ומשנה את הגוון מצהוב או חום ללבן. מבהירים אופטיים מצויים בחומרי ניקוי, כביסה, נייר, קוסמטיקה, ביגוד ועוד.
^Valeur, B.; Berberan-Santos, M. R. N. (2011). "A Brief History of Fluorescence and Phosphorescence before the Emergence of Quantum Theory". Journal of Chemical Education. 88 (6): 731–738. Bibcode:2011JChEd..88..731V. doi:10.1021/ed100182h.
^Muyskens, M.; Ed Vitz (2006). "The Fluorescence of Lignum nephriticum: A Flash Back to the Past and a Simple Demonstration of Natural Substance Fluorescence". Journal of Chemical Education. 83 (5): 765. Bibcode:2006JChEd..83..765M. doi:10.1021/ed083p765.
^Stokes (1852), pages 472–473. In a footnote on page 473, Stokes acknowledges that in 1843, Edmond Becquerel had observed that quinine acid sulfate strongly absorbs ultraviolet radiation (i.e., solar radiation beyond Fraunhofer's H band in the solar spectrum). See: Edmond Becquerel (1843) "Des effets produits sur les corps par les rayons solaires"Archived
^Holler, F. James; Skoog, Douglas A. and Crouch, Stanley R. (2006) Principles Of Instrumental Analysis. Cengage Learning. ISBN 0495012017
^"Animation for the Principle of Fluorescence and UV-Visible Absorbance" Archived 9 June 2013 at the Wayback Machine.. PharmaXChange.info
^ 12Lakowicz, Joseph R. (1999). Principles of Fluorescence Spectroscopy. Kluwer Academic / Plenum Publishers. ISBN 978-0-387-31278-1.
^Valeur, Bernard, Berberan-Santos, Mario (2012). Molecular Fluorescence: Principles and Applications. Wiley-VCH. ISBN 978-3-527-32837-6. p. 64
^Valeur, Bernard, Berberan-Santos, Mario (2012). Molecular Fluorescence: Principles and Applications. Wiley-VCH. ISBN 978-3-527-32837-6. p. 186
^IUPAC. Kasha–Vavilov rule – Compendium of Chemical Terminology, 2nd ed. (the "Gold Book") Archived 21 March 2012 at the Wayback Machine. Compiled by McNaught, A.D. and Wilkinson, A. Blackwell Scientific Publications, Oxford, 1997.
^ 12Wucherer, M. F.; Michiels, N. K. (2012). "A Fluorescent Chromatophore Changes the Level of Fluorescence in a Reef Fish". PLoS ONE. 7 (6): e37913. Bibcode:2012PLoSO...737913W. doi:10.1371/journal.pone.0037913. PMC 3368913. PMID22701587.
^Fujii, R (2000). "The regulation of motile activity in fish chromatophores". Pigment Cell Research. 13 (5): 300–19. doi:10.1034/j.1600-0749.2000.130502.x. PMID11041206.
^ 12Sparks, J. S.; Schelly, R. C.; Smith, W. L.; Davis, M. P.; Tchernov, D.; Pieribone, V. A.; Gruber, D. F. (2014). Fontaneto, Diego (ed.). "The Covert World of Fish Biofluorescence: A Phylogenetically Widespread and Phenotypically Variable Phenomenon". PLoS ONE. 9 (1): e83259. Bibcode:2014PLoSO...983259S. doi:10.1371/journal.pone.0083259. PMC 3885428. PMID24421880.
^Mazel, Charles (2017). "Method for Determining the Contribution of Fluorescence to an Optical Signature, with Implications for Postulating a Visual Function". Frontiers in Marine Science. 4. doi:10.3389/fmars.2017.00266. ISSN 2296-7745.
^Matz, M. "Fluorescence: The Secret Color of the Deep". Office of Ocean Exploration and Research, U.S. National Oceanic and Atmospheric Administration. Archived from the original on 31 October 2014.
^Heinermann, P (10 March 2014). "Yellow intraocular filters in fishes". Experimental Biology. 43 (2): 127–147. PMID6398222.
^Bou-Abdallah, F.; Chasteen, N. D.; Lesser, M. P. (2006). "Quenching of superoxide radicals by green fluorescent protein". Biochimica et Biophysica Acta (BBA) - General Subjects. 1760 (11): 1690–1695. doi:10.1016/j.bbagen.2006.08.014. PMC 1764454. PMID17023114
^Field, S. F.; Bulina, M. Y.; Kelmanson, I. V.; Bielawski, J. P.; Matz, M. V. (2006). "Adaptive Evolution of Multicolored Fluorescent Proteins in Reef-Building Corals". Journal of Molecular Evolution. 62 (3): 332–339. Bibcode:2006JMolE..62..332F. doi:10.1007/s00239-005-0129-9. PMID16474984
^Andrews, K; Reed, S. M.; Masta, S. E. (2007). "Spiders fluoresce variably across many taxa". Biology Letters. 3 (3): 265–7. doi:10.1098/rsbl.2007.0016. PMC 2104643. PMID17412670.
^Stachel, S. J.; Stockwell, S. A.; Van Vranken, D. L. (1999). "The fluorescence of scorpions and cataractogenesis". Chemistry & Biology. 6 (8): 531–539. doi:10.1016/S1074-5521(99)80085-4. PMID10421760.
^Iriel, A. A.; Lagorio, M. A. G. (2010). "Is the flower fluorescence relevant in biocommunication?". Naturwissenschaften. 97 (10): 915–924. Bibcode:2010NW.....97..915I. doi:10.1007/s00114-010-0709-4. PMID20811871
^McDonald, Maurice S. (2 June 2003). Photobiology of Higher Plants. John Wiley & Sons. ISBN 9780470855232.
^Harris, Tom (7 December 2001). "How Fluorescent Lamps Work". HowStuffWorks. Discovery Communications. Archived from the original on 6 July 2010. Retrieved 27 June 2010.
^Rye, H. S.; Dabora, J. M.; Quesada, M. A.; Mathies, R. A.; Glazer, A. N. (1993). "Fluorometric Assay Using Dimeric Dyes for Double- and Single-Stranded DNA and RNA with Picogram Sensitivity". Analytical Biochemistry. 208 (1): 144–150. doi:10.1006/abio.1993.1020. PMID7679561.
^Hawkins, H. Gene; Carlson, Paul John and Elmquist, Michael (2000) "Evaluation of fluorescent orange signs" Archived 4 March 2016 at the Wayback Machine, Texas Transportation Institute Report 2962-S.





 ערך מורחב – פוטוכימיה
ערך מורחב – פוטוכימיה












![{\displaystyle \left[S_{1}\right]=\left[S_{1}\right]_{0}e^{-\Gamma t}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/9862f745e8c8e8f83083c7e8038c0a4c632b6c07)
![{\displaystyle \left[S_{1}\right]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/85875c6a1407cb88df37cff6cac722a1b488dbc2)

![{\displaystyle \left[S_{1}\right]_{0}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/8ddfd576e02a185cecd193db9b729e228db24d84)




















,_Monterey_Bay_Aquarium,_Monterey,_California,_USA.jpg)










_B2.jpg)

 קרינה פלואורסצנטית, דף שער בספרייה הלאומית
קרינה פלואורסצנטית, דף שער בספרייה הלאומית גרדיאן, רבים מהיונקים בטבע זוהרים באור אולטרה־סגול, מסיבה שטרם התבררה, באתר הארץ, 5 באוקטובר 2023
גרדיאן, רבים מהיונקים בטבע זוהרים באור אולטרה־סגול, מסיבה שטרם התבררה, באתר הארץ, 5 באוקטובר 2023