אבולוציה של האדם
הָאֵבוֹלוּצְיָה שֶׁל הָאָדָם היא תהליך ההתפתחות (אבולוציה) של המין האנושי המודרני, אדם נבון (Homo sapiens), כמו גם של מינים נכחדים נוספים בתת-שבט האדם, מתוך קופי אדם קדומים. תהליך זה התרחש במשך כעשרת מיליוני השנים האחרונות, במהלך התקופות הגאולוגיות של המִיוֹקֵן, הפְּלִיוֹקֵן, הפְּלֶייסטוֹקֵן וההוֹלוֹקֵן. האבולוציה של האדם מוכרת למדע מתוך השוואת האנטומיה, ההתנהגות ורצפי הגנום של האדם ושל קופי האדם, מתוך חקירת מאובנים של מינים נכחדים ומתוך שרידים ארכאולוגיים כגון כלי אבן מסותתים ואמנות פרהיסטורית. ממצאי המאובנים חושפים את קיומה של שושלת מסועפת, שבה לפחות עשרים מינים ידועים, שמתוכם שרדו עד ימינו רק האדם ומספר קופי אדם, כמו השימפנזה והגורילה. על פי ממצאים אלו, התהליכים העיקריים שהובילו להתפתחות האדם הם ההזדקפות על הרגליים האחוריות, שהחלה לפני כשבעה עד חמישה מיליוני שנים, ולאחריה הגידול המהיר של קליפת המוח, בשיעור של בערך פי שלושה בתוך שני מיליוני השנים האחרונות. על פי תורת האבולוציה המודרנית, תהליך זה התרחש כתוצאה ממוטציות ומגוון גנטי שנבררו על ידי ברירה טבעית וסחף גנטי.
האבולוציה של האדם היא נושא למחקר לא רק עבור העוסקים במדעי החיים אלא גם עבור אלו העוסקים בקשת רחבה של מדעי החברה ומדעי הרוח. תחום הפלאואנתרופולוגיה מאחד את חקר המאובנים עם מחקר אנתרופולוגי של חברות אדם עתיקות, ותחום הארכאולוגיה הפרהיסטורית חוקר את התרבות החומרית שלהן. תחומי הגנטיקה האבולוציונית, הגנומיקה והביואינפורמטיקה עוסקים בין השאר בפענוח האבולוציה של האדם על פי רצפי דנ"א ורנ"א, הפרימטולוגיה עוסקת בחקר הביולוגיה וההתנהגות של קופים וקופי אדם בני ימינו, והפסיכולוגיה האבולוציונית חוקרת את ההתנהגות של בני אדם בני ימינו מנקודת מבט אבולוציונית.
היסטוריה של חקר מוצא האדם עריכה
 ערך מורחב – היסטוריה של חקר מוצא האדם
ערך מורחב – היסטוריה של חקר מוצא האדם
במאה ה-18 פתח חוקר הטבע השוודי קארולוס לינאוס במבצע מיון טקסונומי של כל עולם החי והצומח על בסיס המורפולוגיה והאנטומיה שלהם. את האדם והקופים הוא סיווג לאותה סדרה – ה"פרימטים" (מלטינית "ראשונים במעלה"), ולאדם העניק את השם המדעי בלטינית "הוֹמוֹ סָפִּיֶינְס" (Homo sapiens) – שם הסוג "הומו" פירושו "אדם", ושם המין "ספיינס" פירושו "נבון". מיון עולם החי חשף את הסדר ההיררכי השורר בו, ובתחילת המאה ה-19 החלו חוקרי טבע כמו ז'אן-בטיסט דה לאמארק לבדוק את רעיון המוצא המשותף של המינים על מנת להסבירו. בשנת 1859 פרסם צ'ארלס דרווין את המהדורה הראשונה של ספרו "מוצא המינים", שהציע מנגנון של שׁוֹנוּת וברירה טבעית. בספר זה נמנע דרווין מלהתייחס לסוגיה הטעונה של מוצא האדם, אבל תומאס הקסלי עשה זאת לראשונה במאמר ההיסטורי "ראיות למקומו של האדם בטבע" ב-1863, וב-1871 פרסם דרווין את ספרו "מוצא האדם", שבו חזה כי יתגלו מאובני מעבר: צורות ביניים בין קופי האדם לאדם, השערה שמולה עמדה השערה נגדית שחזתה שככל שיימצאו מאובנים נוספים הם יהיו דומים למינים המוכרים ולא יתכנסו אל צורות הביניים[1].
התחום המדעי המתרכז במאובני משפחת האדם נקרא פלאואנתרופולוגיה. בימיו של דרווין כבר התגלו באירופה כמה שלדים מאובנים של אדם ניאנדרטלי, אבל לא היה ברור אם הם מין נפרד מן האדם. דוגמה מובהקת ראשונה של בני אדם קדומים בעל מוח הקטן משמעותית ממוח אדם מודרני הייתה מאובן הומו ארקטוס שהתגלה בשנת 1891 באי ג'אווה שבדרום-מזרח אסיה. התפתחויות חשובות בתחום זה במאה ה-20 היו גילוי מאובני האוסטרלופיתקנים ומינים קדומים של סוג האדם במזרח אפריקה ודרומה, על ידי פלאואנתרופולוגים כריימונד דארט, רוברט ברום, משפחת ליקי, דונלד ג'והנסון וטים וייט. תגליות אלו ביססו את מוצאה האפריקני של משפחת האדם, ואת העלייה ההדרגתית שחלה בגודל המוח במשך שלושת מיליוני השנים האחרונות. עד היום נתגלו כמה אלפי מאובנים ממשפחת האדם, המסווגים כיום ל-9 עד 24 מינים נכחדים של הומינינים מדרגות שונות של שלבי ביניים בין קופי האדם לאדם.
 |
מאז שנות ה-60 של המאה ה-20 מתחוללת מהפכה בתחום גילוי המאובנים הודות לשיטות תִארוך רדיומטריות ופלאומגנטיות, מהן עולה שהאבולוציה של האדם התרחשה במשך שבעת מיליוני השנים האחרונות לכל הפחות, במהלך התקופות הגאולוגיות של המיוקן, הפליוקן, הפלייסטוקן וההולוקן. עדויות ארכאולוגיות למוצא האדם, בעיקר של כלי-אבן מסותתים, נפוצות הרבה יותר מאשר עדויות מאובנים, וכיום מוכרות היטב מספר תרבויות ייצור כלים מן התקופה שבמינוח הארכאולוגי נקראת התקופה הפלאוליתית. במקביל התפתחו במאה ה-20 תחומי המחקר הפרימטולוגי של קופים והמחקר האנתרופולוגי של תרבויות ציידים-לקטים בימינו, תחומים התורמים להבנה טובה יותר של ממצאי המאובנים.
מהפכה נוספת בחקר האבולוציה של האדם מתרחשת הודות לפענוח הקוד הגנטי ומבנה מולקולת הדנ"א האחראית לתורשה של יצורים חיים. החל משנות ה-70 של המאה ה-20 התאפשר מיפוי הרצף של גנים רבים, עד כדי מיפוי גנום האדם כולו, וכן גנום השימפנזה[2], בתחילת המאה ה-21. בתחום הגנטיקה האבולוציונית האנושית, שיטות שעון מולקולרי תארכו את האב הקדמון המשותף של האדם ושל השימפנזה לתקופה של 8 עד 4 מיליון שנים לפני זמננו, והראו שמוצא האנושות בת-ימינו הוא מאוכלוסיית בסיס קטנה שהיגרה מאפריקה במהלך כ-200 אלף השנים האחרונות. גנטיקאים אבולוציוניים כמו סוונטה פבו פיתחו שיטות פלאוגנטיות למיצוי דנ"א מעצמות מאובנות, שאפשרו גם את מיפוי הגנום הניאנדרטלי, שעל פיו מחשבים כי האב הקדמון המשותף שלו ושל האדם המודרני התקיים לפני כ-500 אלף שנה. בשנים האחרונות מתחקה המחקר אחר השינויים הגנטיים האחראים לאנושיותו של האדם.
עדויות ביולוגיות למוצא האדם עריכה
מקומו של האדם בטבע עריכה

- ערך מורחב – חיים
מנקודת מבט ביוכימית, אנטומית, פיזיולוגית וגנטית האדם הוא מין של יצור חי, שאינו נבדל באופן עקרוני מכל שאר המינים החיים על פני כדור הארץ. האדם חולק עמם אותן מולקולות בסיסיות של החיים כמו דנ"א, רנ"א, חומצות אמינו, פחמימות ושומנים, אותו קוד גנטי, ואף הגנים שלו דומים מאוד (ולעיתים אף זהים) לגנים של היצורים החיים הקרובים לו. כפי שמצא לינאוס, ניתן לסווג את האדם על פי אותם קריטריונים ששימשו לסיווג שאר עולם החי. מבחינת סיווגו הטקסונומי האדם הוא:
- איקריוט: התאים שלו בעלי גרעין ואברונים.
- בעל חיים: הוא מורכב ממספר טיפוסי תאים מתמחים כולל תאי עצב.
- מיתרן: יש לו מיתר לאורך הגב, כיסי זימים בגרון וזנב (בשלב העוברי).
- חולייתן: יש לו שלד פנימי.
- יונק: הוא אנדותרמי, בעל שיער והוולדות שלו יונקים חלב מן האם.
- יונק שליה: הוא יולד ולדות חיים הניזונים ברחם האם באמצעות שליה.
- פרימט: הוא בעל ידיים עם חמש אצבעות ארוכות, ציפורניים במקום טפרים, מוח גדול יחסית לגופו, אף קטן, עיניים בעמדה קדמית, ונוסחת שיניים של שתי חותכות, ניב אחד, שתי קדם-טוחנות ושלוש טוחנות בכל רבע לסת.
על פי הדמיון ברצף הדנ"א בגנום ניתן כיום לשרטט בדיוק רב את שושלת המינים החיים ובתוכם האדם.
מצד שני, האדם הוא הפרימט בעל המוח הגדול ביותר ביחס למשקל גופו[3], ועם מוח גדול משמעותית מהמצופה מיצור בגודלו של האדם[4]. מגמת גידול המוח ניכרת כבר באבולוציה של החולייתנים, היונקים, הפרימטים וקופי האדם, אך מוח האדם גדול פי שלושה לערך מזה של קופי האדם החיים כיום. במדע המודרני מקובל שהבדל זה בגודל המוח הוא העומד ביסוד ייחודו הקוגניטיבי, הנפשי והתרבותי של האדם[3]. תכונות אנטומיות נוספות ייחודיות של האדם הן גפיים קדמיות (ידיים) בעלות אצבעות גמישות ורגישות ואגודל נגדי, המאפשרים טיפול עדין בעצמים וכלים, יציבה זקופה המשחררת את הידיים ממשימת ההתקדמות, וגרון עמוק המאפשר יכולת משוכללת של דיבור. כמו כן, האדם הוא היונק בעל תקופת הינקות וההתבגרות הממושכת ביותר, יחסית לתוחלת החיים הכללית שלו.
מוצא הפרימטים, הקופים וקופי האדם עריכה


- ערך מורחב – פרימטים
סדרת הפרימטים מופיעה ברשומות המאובנים לפחות מאז תקופת האאוקן, לפני כ-50 מיליון שנה, ועל פי תארוכים גנומיים ייתכן שחבריה הראשונים אף קדמו להכחדת הדינוזאורים בסוף תור הקרטיקון. מכל מקום הפרימטים של האאוקן כבר טיפוסיים מאוד לצורה המודרנית של בני הסדרה, עד כי אין כל קושי בזיהויים[5]. הפרימטים הם יונקים שוכני עצים ביערות טרופיים וסובטרופיים, בעלי גפיים ארוכות וגמישות עם חמש אצבעות ארוכות בכל גף, שבקצותיהן כריות אצבע רגישות וציפורניים שטוחות במקום טפרים. הגפיים והאצבעות הם בבירור הסתגלות ללפיתת ענפים, אך בדיעבד הסתברו כיעילים מאוד לטיפול עדין בכלים. בדומה לכך, לפרימטים עיניים גדולות שפונות קדימה, ולא לצדדים כמו במרבית היונקים, ומאפשרות להם ראייה סטראוסקופית ואומדן מרחקים מדויק. העיניים האלה הן בבירור הסתגלות לתנועה בזינוקים בין ענפים, אך בדיעבד הסתברו כיעילות לצורך בחינה מדוקדקת של עצמים וטיפול בכלים. תופעה כזו של הסתגלות, שבדיעבד מסתברת כמתאימה למטרה אחרת, היא תופעה נפוצה בהתפתחות אבולוציונית ומכונה פרה-אדפטציה. מבין התכונות האחרות האופייניות לפרימטים ניתן למנות ראיית צבע מעולה, חוש ריח חלש יחסית המתבטא באונות ריח קטנות וזרבובית קטנה יחסית לשאר היונקים, ומספר קטן של צאצאים בהמלטה, לרוב לא יותר מאחד או שניים שאותם האם נושאת איתה. בהתאם לכך לפרימטים יש זוג פטמות יחיד הממוקם על החזה.
- ערך מורחב – קופים
סדרת הפרימטים כוללת מספר תת-סדרות קיימות ונכחדות, שהגדולה והמפותחת מתוכן היא תת-סדרת הקופים (Infraorder: Simiiformes). הקופים הם פרימטים פעילי-יום, בעלי מוח גדול יחסית לגופם, אינטליגנציה גבוהה, התנהגות חברתית מפותחת, תקופת התבגרות ארוכה של הצעירים, ויכולת ללמוד באמצעות חיקוי ההתנהגות של פרטים אחרים. במשך הפלאוקן והאאוקן יבשות גונדוונה ולאוראסיה התפצלו אט-אט ליבשות המודרניות, וכתוצאה מכך התפתחה הפרדה בין הקופים "רחבי האף" (Platyrrhini), שהפכו למבודדים בדרום אמריקה, לבין הקופים "צרי האף" (Catarrhini) בעולם הישן. האחרונים הם הקבוצה הגדולה והמגוונת יותר, אך את כולם, לרבות האדם, מאפיינת נוסחת שיניים של שתי חותכות, ניב אחד, שתי קדם-טוחנות (מלתעות) ושלוש טוחנות בכל רבע לסת, ובסך הכל 32 שיניים בעלות תצורה בלתי-מתמחה, כלומר הן אינן מותאמות באופן מיוחד לטריפה מצד אחד או לגריסת מזון צמחי קשה מצד אחר, אלא לתפריט מגוון של אוכלי-כול[5].
בתקופת האוליגוקן הקופים צרי-האף התפצלו לשתי על-משפחות: קופי העולם הישן בעלי הזנב (Superfamily: Cercopithecoidea) וקופי האדם (Superfamily: Hominoidea). קופי האדם הם באופן כללי בעלי גוף גדול יותר, והזנב שלהם, שבשאר קבוצות הפרימטים הוא לרוב מפותח, התנוון[6]. הם היו בתחילה הקבוצה המצליחה יותר, ומתקופת המיוקן ידועים עשרות מינים מאובנים שלהם על פני אסיה, אירופה ואפריקה. אבל במהלך הפליוקן והפלייסטוקן התהפך הגלגל, וכל קופי האדם פרט לסוג האדם שרדו רק ביערות גשם טרופיים באפריקה ובמזרח אסיה, אולי כתוצאה מתחרות עם הקופים בעלי הזנב, שכיום מיוצגים על ידי סוגים מצליחים ומרובי מינים כמו המקוק, הגנון והבבון. כיום קופי האדם מחולקים לשתי משפחות: הגיבוניים (Family: Hylobatidae) וקופי האדם הגדולים, או ההומינידיים (Family: Hominidae), שממנה שרדו כיום האדם, שני מיני שימפנזה, שני מיני גורילה ושני מיני אורנגאוטן[5].

- ערך מורחב – ניסויים בלשניים בקופי אדם
קופי האדם הגדולים, ובעיקר מיני השימפנזה, הם בין בעלי-החיים הנבונים ביותר. שימפנזים לומדים לזהות את עצמם במראה, יכולת שחוקרים מסוימים מחשיבים כראיה למודעות עצמית[7], והודגם כי במידה מסוימת יש להם תאוריה של הנפש, כלומר הם מייחסים כוונות ומצבים נפשיים לקופים אחרים[8][9]. הם מסוגלים לפתור בעיות המצריכות דמיון והסקה, שחלקן קשות אפילו לבני אדם. במקום חיותם בטבע שימפנזים משתמשים לעיתים במגוון כלים שונים, כגון מקלות לשליית טרמיטים ואבנים לפיצוח אגוזים. הם בוחרים כלים כאלו בשימת לב[10], ואף מתאימים אותם לשימוש הרצוי באופנים פשוטים, למשל שבירת ענף ליצירת מקל חד לצורך ציד חיות קטנות בתוך נקרות עץ[11]. מכיוון שהם לומדים כישורים כאלו על ידי חיקוי שימפנזים אחרים, נוצרות אצלם בטבע "תרבויות" האופייניות לכל קהילה ושונות במנהגיהן מאלו של קהילות אחרות[12]. סביר להניח שכישורים דומים התקיימו באב הקדמון המשותף של האדם והשימפנזה. בשִׁבְיָה שימפנזים מסוגלים ללמוד להשתמש בכלים רבים של בני-אדם, ואף לסתת כלי-אבן פשוטים[13]. הם אינם מסוגלים לדבר משום שהאנטומיה של הגרון שלהם אינה מתאימה לכך, אך הם יכולים ללמוד להבין עשרות ומאות מילים בשפה אנושית, לתקשר באופן בסיסי בשפות סימבוליות שאינן דורשות דיבור, כמו שפת סימנים של לקויי שמיעה, ואף להמציא צירופי מילים משל עצמם, ברמה שתוארה כדומה לזו של ילדים אנושיים בני שנתיים וחצי[14]. ישנו ויכוח בין חוקרים בתחום הבלשנות עד כמה יכולות אלו משתוות לשפה אמיתית. שימפנזים גם למדו כישורים מתמטיים בסיסיים כמו מנייה וחיבור[15], כולל מושג האפס[16], ושימוש בקטגוריות מופשטות (למשל "זהה" ו"שונה").
ההומינידים: משפחת האדם עריכה
- ערך מורחב – הומינינים


הסיווג של האדם וקופי האדם הגדולים עבר שינויים רבים בחצי המאה האחרונה. בתחילת שנות ה-60 של המאה ה-20 עדיין היה מקובל לייחד את משפחת ההומינידים אך ורק לאדם ולאבותיו הנכחדים, בעוד קופי האדם הגדולים הופרדו למשפחה אחרת אם כי קרובה, הפונגידיים (Pongidae על שמו של Pongo, השם המדעי של האורנגאוטן). הדעה השלטת באותה תקופה ראתה בפרוקונסול, קוף אדם מאובן מלפני כ-20 מיליון שנה, את האב הקדמון המשותף של האדם והשימפנזה. אך כתוצאה מן ההתקדמות בחקר הגנום, הוברר שהאב הקדמון המשותף חי בתקופה הרבה יותר מאוחרת, בין 4 ל-8 מיליון שנה, כלומר השימפנזה קרוב לאדם הרבה יותר ממה שהיה מקובל לחשוב ואין כל הצדקה להפרידו למשפחה אחרת. יתר על כן, המחקר הגנטי הראה שהשימפנזה קרוב לאדם יותר מאשר לגורילה, ושהגורילה קרוב לאדם ולשימפנזה יותר מאשר לאורנגאוטן. מכך נובע שכל יחידה טקסונומית שתכלול את הסוגים שימפנזה, גורילה ואורנגאוטן, אבל לא את האדם עצמו תהיה בהכרח פרפילטית – היא תכלול שלושה ענפים נפרדים, בניגוד למוסכמה לכלול בכל יחידה טקסונומית ענף מונופילטי יחיד[17].
התוצאה הסופית של תסבוכת זו הייתה שמשפחת הפונגידיים בוטלה לחלוטין, וקופי האדם הגדולים הוכנסו אחר כבוד למשפחת ההומינידיים עם האדם. על מנת לייחד את ענף האדם מענף השימפנזה, נדרש אפוא להוסיף רמה טקסונומית נוספת מתחת רמת המשפחה ומעל רמת הסוג – ה"שבט" (tribe). הטקסונומים עדיין חלוקים בדעותיהם לגבי הסיווג הראוי ברמת השבט ותת-השבט, אך כולם מייחדים כעת לענף האדם את הכינוי "הומינינים" (hominins או homininans)[17]. למרות (או אולי בגלל) תסבוכת זו, בשפה לא-רשמית אפילו פלאואנתרופולוגים רבים עדיין משתמשים במונח "הומינידים" (hominids) לציון האדם וקרוביו הנכחדים השייכים לאותו ענף, ובמונח "קופי אדם" לציון שאר הענפים.
קרבתו הגנטית של האדם לשאר הפרימטים עריכה

- ערך מורחב – גנטיקה אבולוציונית אנושית
שיעור ההבדל בין גנום האדם לגנום השימפנזה, כשהוא נמדד במספר זוגות הנוקלאוטידים השונים מתוך כלל הגנום, הוא כאחוז אחד[2]. למעשה מחקרים שונים נוקבים באחוזים מעט שונים כהבדל בין גנום האדם והשימפנזה, והסיבה לכך היא השימוש במדגמים שונים ודרכים שונות לחישוב הדמיון. אך בכל שיטה בה משתמשים לאומדן, ההבדלים בין בני אדם לשימפנזים גדולים רק פי עשרה לערך מן ההבדלים בין בני אדם שונים. בהנחת קצב קבוע של הצטברות מוטציות (הנחת "השעון המולקולרי"), ניתן להסיק שהזמן שחלף מאז האב הקדמון המשותף לאדם ולשימפנזה ארוך רק בסדר גודל אחד מהזמן שחלף מאז האב הקדמון של כל בני האדם החיים כיום. הנחה זו מבוססת על התגליות בגנטיקה של אוכלוסיות כי מרבית המוטציות הן נייטרליות, ומתקבעות באוכלוסייה בסחף גנטי. מדידות גנטיות מעריכות את קצב ההופעה של מוטציות חדשות בסדר גודל של 100 לגנום לדור, ומכך ניתן להעריך שהאב הקדמון של האדם והשימפנזה חי לפני מספר מיליוני שנים. בפועל שיטות שעון מולקולרי הן מתוחכמות יותר, ומתחשבות גם בגודל האוכלוסייה, אך כל המחקרים שנערכו בנושא זה בשנים האחרונות נותנים הערכות של בין 4 מיליון ל-8 מיליון שנה[18][19]. ההערכה המעודכנת לגילה של חוה המיטוכונדרית – האֵם הקדמונית המשותפת המאוחרת ביותר של כל בני האדם החיים כיום דרך שושלות אימהיות בלבד – היא כ-170 אלף שנה[20]. פענוח הגנום הניאנדרטלי על ידי מיצוי דנ"א מעצמות מאובנות אִפשר להוסיף את הניאנדרטלים לחישובים אלו. גיל האב הקדמון המשותף לאדם המודרני ולניאנדרטלים מוערך ב-460 אלף עד 510 אלף שנה[20][21].
ראיות מרשימות ביותר לקרבת המשפחה בין האדם וקופי האדם מודגמות דווקא ברצפי דנ"א שאין להם תפקוד מעשי, אלא הם תוצאה של "תאונות" גנטיות. ניתן להמשיל מצב זה לשני תלמידים שהואשמו בהעתקה במבחן בגלל הדמיון הרב בין תשובותיהם. התלמידים עשויים לתרץ את הדמיון בטענה שהם חשבו באופן בלתי-תלוי על אותה תשובה נכונה, אבל אם מתגלות בתשובות שלהם אפילו שגיאות כתיב זהות, הראיה להעתקה הופכת משכנעת ביותר. שגיאות מעין אלו אכן שכיחות בגנום, למשל פסאודוגנים. דוגמה לכך היא הגן של גולונולאקטון אוקסידאז, האנזים האחרון בשרשרת הייצור של ויטמין C בגוף. גן זה קיים ותקין במרבית החולייתנים, אבל בכל מיני הקופים שנבדקו עד היום, כולל האדם, הוא אינו מצליח להתבטא עקב מוטציה של הזזת מסגרת הקריאה. כתוצאה מפגם גנטי זה האדם והקופים אינם יכולים לייצר ויטמין C בעצמם, והם יחלו ואף ימותו מצפדינה אלא אם כן יקבלו אותו במזונם. את הדמיון הגנטי בין הפסאודוגנים של הקופים והאדם לא ניתן לתרץ באותו התפקוד, וההסבר ההגיוני היחיד הוא שהם ירשו אותה מוטציה שהתרחשה באב הקדמון המשותף שלהם לפני עשרות מיליוני שנה[22]. מוטציה כזו הייתה עשויה להתפשט ולהתקבע באוכלוסיית האב הקדמון כתוצאה מסחף גנטי אם היא לא הייתה מזיקה באותו זמן, למשל אם אותו אב קדמון ניזון מפירות עשירים בוויטמין C ולכן לא נזקק לייצר אותו בעצמו. דוגמה אחרת לשגיאות זהות התגלתה בפסאודוגן נוסף: אחד העותקים של הגן להמוגלובין. עותק זה נמצא במיקום זהה בגנום האדם, בגנום השימפנזה ובגנום הגורילה, אך הוא אינו מסוגל להתבטא כתוצאה ממוטציות התמרה של שני נוקלאוטידים בלבד בתחילת הרצף שלו. שני הנוקלאוטידים המותמרים זהים במדויק בפסאודוגן של האדם, השימפנזה והגורילה. שאר הרצף של הגן הוא כמעט בלתי פגוע וזהה לעותקים אחרים של המוגלובין, אך בהמשכו חסר נוקלאוטיד יחיד, ואף החוסר הזה זהה במדויק בפסאודוגנים של האדם, השימפנזה והגורילה[23].
ייחודו הגנטי של האדם משאר הפרימטים עריכה

גנום האדם וגנום השימפנזה נבדלים זה מזה בכמה עשרות מיליוני נוקלאוטידים. רובם מייצגים מוטציות נייטרליות, שאינן תורמות להבדלים הפנוטיפיים בין האדם ובין השימפנזה. קשה מאוד לגלות את הנוקלאוטידים המעטים מתוכם שכן תורמים להבדלים הפנוטיפיים, כלומר, את השינויים הגנטיים האחראים לאנושיותו של האדם. אף על פי שהגנומים של שני המינים כבר מופו, לא ידוע עדיין על אף גן מקודד לחלבון באדם שאינו קיים ברצף זהה או כמעט זהה בשימפנזה. לכן סביר להניח כי ההבדל טמון ברצפי בקרה – רצפי דנ"א המבקרים את פעולתם של גנים מקודדים לחלבונים. אחת השיטות לגלות רצפים כאלו היא לחפש אזורים בגנום שלא השתנו הרבה בחולייתנים קדומים ובקו ההתפתחות של השימפנזה, ולעומת זאת עברו אבולוציה מואצת בקו ההתפתחות של האדם. אזורים כאלה קיבלו את הכינוי HAR (ראשי תיבות של Human Accelerated Regions). כמאתיים מהם מופו בשנת 2006 במחקר של קתרין פולארד ועמיתים מאוניברסיטת קליפורניה בסנטה קרוז[24]. למשל HAR1, בעל האבולוציה המהירה מבין אלו שהתגלו, הוא מקטע על כרומוזום 20 בן 118 זוגות נוקלאוטידים, ש-18 מתוכם השתנו בענף האדם מאז התפצלותו מענף השימפנזה, לפני כ-5 מיליון שנים, לעומת 2 זוגות נוקלאוטידים בלבד שהשתנו במשך 300 מיליון השנים שלפני כן, מאז האב הקדמון המשותף לאדם, לשימפנזה ולתרנגולת. הסתבר שקיימת חפיפה חלקית בין HAR1 ובין שני גנים של רנ"א שאינם מתורגמים לחלבון אלא מתועתקים למולקולות רנ"א שליח המשפיעות על התעתוק של גנים אחרים. התגלה כי לאחד משני הגנים האלו, HAR1F, יש תפקיד בהתפתחות הרקמה העוברית של תאי עצב בקליפת המוח[25], אף כי אופן פעולתו המדויק עדיין אינו ידוע.
אזור HAR מפורסם אחר הוא הגן FOXP2 על כרומוזום 7 המקודד לגורם שעתוק. מוטציות שחלו בו בשני נוקלאוטידים משותפות לגנום האדם ולגנום הניאנדרטלי, אך לא לגנום השימפנזה ושאר החולייתנים, והן קשורות ככל הנראה ביכולת הדיבור. הכנסת שתי המוטציות האלו לגן FOXP2 של עכברים גרמה לשינויים בתאי עצב במוח הגדול ולשינויים בקולות שהם משמיעים[26].
תאוריה שהוצעה כבר במחצית המאה ה-20, בין השאר על ידי מדענים ידועי-שם כאשלי מונטגיו וסטיבן גולד, גורסת שהאבולוציה של האדם היא במידה רבה תוצאה של נאוטניה. נאוטניה במספר גדול של מערכות בגוף עשויה להיגרם כתוצאה משינויים במספר קטן של רצפי בקרה שמפעילים את אותם גנים, ולכן תאוריה כזו עשויה להסביר את האבולוציה המקיפה שחלה באדם תוך מיליוני שנה בודדים. לפי תאוריה זו, האדם שונה משאר קופי האדם משום שהוא שומר גם בבגרותו על תכונות של קופי אדם צעירים. בפרט הוצע שהגידול במוח האדם הוא תוצאה של הארכת השלב בינקות שבו המוח עושה חלק ניכר מגידולו. מחקרים מהשנים האחרונות מראים שגנים רבים במוח האנושי אכן מתבטאים בשלב מאוחר יותר של חיי הפרט, יחסית למוחות שימפנזים[27]. עם זאת, מחקרים אלו מראים גם שמספר קטן יותר של גנים אחרים מתבטאים דווקא בשלב מוקדם יותר. מסיבה זו חוקרים כיום מעדיפים להשתמש במונח "הטרוכרוניות" (heterochrony), כלומר אבולוציה כתוצאה משינוי מועדי התבטאות של אותם גנים בהתפתחות העוברית והתבגרות הפרט[3].
עדות המאובנים לאבולוציה של האדם עריכה
- ערך מורחב – תיארוך רדיומטרי
מאז התפצלות ההומינינים מענף השימפנזה לפני מספר מיליוני שנים ועד להתפצלות של ענף האדם הניאנדרטלי לפני כחצי מיליון שנה, לא שרד גנום אחר כלשהו שניתן להשוותו לגנום האדם בן-ימינו. לכן, למרות החידושים מתחום הגנטיקה האבולוציונית, גם כיום חקר האבולוציה של האדם במשך תקופה זו מסתמך בעיקר על גילויים של שרידים מאובנים. חקר המאובנים מראה על קיומה של שושלת מסועפת שממנה מוכרים כיום 9 עד 24 (תלוי באופן הסיווג) מינים ידועים של הומינינים. הסיווג נעשה באמצעות תכונות אנטומיות, כמו גודל המוח, הניתן למדידה על ידי הנפח הפנימי של חלל הגולגולת, ומורפולוגיית השלד והשיניים. את מרבית המאובנים ניתן לתארך באמצעות מגוון שיטות תארוך פיזיקליות, ולכן ניתן להדגים שתכונות אלו התפתחו באופן רציף במשך תקופה ארוכה. אך בשל ריבוי המינים, הדמיון הרב ביניהם וצורתו המסועפת של עץ המשפחה של האדם, קשה מאוד לקבוע במדויק איזה מין התפתח מאיזה מין אחר. במשך תקופות ארוכות באבולוציה של האדם התקיימו מספר מיני הומינינים באותו זמן, ולעיתים אף חיו זה לצד זה, אך גם קיום שני מינים בו-זמנית אינו פוסל את האפשרות שאחד מהם התפצל מהשני.

אבות קדמונים באפריקה של המיוקן והפליוקן עריכה
כל המחקרים הגנומיים בשנים האחרונות מעריכים שהאב הקדמון המשותף האחרון לאדם ולשימפנזה חי באפריקה בסוף עידן המיוקן ובתחילת עידן הפליוקן, כ-8 מיליון עד 4 מיליון שנה לפני זמננו, ושהאב הקדמון המשותף לאדם, השימפנזה והגורילה קדם לו בכמיליון עד 2 מיליון שנה. ואולם במשך זמן רב לא ניתן היה לקבל אישוש לכך ברשומות המאובנים, משום שלא התגלה באפריקה אף מאובן של קוף אדם או הומינין מן התקופה שבין 13 מיליון ל-4 מיליון שנה לפני זמננו. חוקרים אחדים אף הציעו כמועמדים חלופיים כמה מקופי האדם שהיו נפוצים בתקופה זו בדרום אירופה, כמו דריופיתקוס מצרפת ואורנופיתקוס מיוון. אולם נראה כי הסיבה להיעדר הממצאים הייתה מלאכותית ונעוצה בעיקר במיעוט מחקרים לחיפוש מאובנים באפריקה יחסית לאירופה. מכל מקום, בשנים האחרונות פורסמו מספר מאובנים אפריקניים חשובים מהתקופה המדוברת, אף כי הם עדיין נדירים מאוד.
-
 סהלאנתרופוס צ'אדנסיס, גיל: 6.5 מיליון שנה, נפח מוח: 340 סמ"ק
סהלאנתרופוס צ'אדנסיס, גיל: 6.5 מיליון שנה, נפח מוח: 340 סמ"ק -
 ארדיפיתקוס רמידוס, גיל: 4.4 מיליון שנה, נפח מוח: 325 סמ"ק
ארדיפיתקוס רמידוס, גיל: 4.4 מיליון שנה, נפח מוח: 325 סמ"ק -
 אוסטרלופיתקוס אפרנסיס, גיל: 3.5 מיליון שנה, נפח מוח: 350 סמ"ק
אוסטרלופיתקוס אפרנסיס, גיל: 3.5 מיליון שנה, נפח מוח: 350 סמ"ק -
 פראנתרופוס אתיופיקוס, גיל: 2.6 מיליון שנה, נפח מוח: 410 סמ"ק
פראנתרופוס אתיופיקוס, גיל: 2.6 מיליון שנה, נפח מוח: 410 סמ"ק -
 אוסטרלופיתקוס אפריקנוס, גיל: 2.0 מיליון שנה, נפח מוח: 400 סמ"ק
אוסטרלופיתקוס אפריקנוס, גיל: 2.0 מיליון שנה, נפח מוח: 400 סמ"ק -
 פראנתרופוס רובוסטוס, גיל: 2 מיליון שנה, נפח מוח: 470 סמ"ק
פראנתרופוס רובוסטוס, גיל: 2 מיליון שנה, נפח מוח: 470 סמ"ק -
 אוסטרלופיתקוס סדיבה, גיל: 1.9 מיליון שנה, נפח מוח: 435 סמ"ק
אוסטרלופיתקוס סדיבה, גיל: 1.9 מיליון שנה, נפח מוח: 435 סמ"ק -
 פראנתרופוס בויזאיי, גיל: 1.7 מיליון שנה, נפח מוח: 525 סמ"ק
פראנתרופוס בויזאיי, גיל: 1.7 מיליון שנה, נפח מוח: 525 סמ"ק



.JPG)



.JPG)
שיני קוף אדם שהתגלו בקניה ב-2007 קיבלו את השם נאקאליפיתקוס Nakalipithecus, והן מתוארכות לגיל של כ-10 מיליון שנה[28]. מסוף המיוקן ותחילת הפליוקן התגלו באפריקה ארבעה מינים: סהלאנתרופוס צ'אדנזיס מצ'אד מלפני 6 מיליון עד 7 מיליון שנה[29], אורורין טוגננסיס מקניה מלפני כ-6 מיליון שנה[30], ושני מינים של ארדיפיתקוס מאתיופיה – ארדיפיתקוס קדבה מלפני 5.5 מיליון שנה[31] וארדיפיתקוס רמידוס[32] מלפני 4.4 מיליון שנה. אך פרט לרמידוס כל הממצאים האלו חלקיים מאוד, ולכן לא ברור עדיין טיב היחסים ביניהם, מי מהם קופי אדם, מי מהם הומינינים ומי מהם מייצג את הפיצול לענפי הגורילה והשימפנזה, אם בכלל. סביר גם שהתקיימו באותה תקופה מינים נוספים בלתי מוכרים, ואם כך ייתכן שהאבות הקדמונים המשותפים לאדם ולקופי האדם האפריקניים עדיין לא התגלו.

ארדיפיתקוס וההזדקפות על שתיים עריכה
- ערך מורחב – ארדיפיתקוס רמידוס
מאובני ארדיפיתקוס רמידוס, ובעיקר שלד הנקבה הידועה בכינוי הפופולרי "ארדי", היא תגלית מבין החשובות ביותר בחקר האבולוציה של האדם בשנים האחרונות. על פי הסברה, ארדי השתייכה לענף ההומיניני מייד לאחר התפצלותו מענף השימפנזה, והיא שופכת לראשונה אור על תהליך ההזדקפות על הרגליים האחוריות[32]. כפות הרגליים של ארדי דומות עדיין לאלו של קופים וקופי אדם – הן בעלות בוהן לופתת נפרדת משאר האצבעות בדומה לאגודל של כף יד – אבל חלקו העליון של האגן שלה הוא כבר ב"צורת קערה", בדומה לאגן האנושי המותאם להליכה על שתיים. שחזור השלד הוליך למסקנה כי ארדי נעה בשתי צורות שונות: בצמרות העצים היא טיפסה על ארבע כפותיה ומעל הענף, בדומה לקופי מקוק ובבון בני-ימינו ובניגוד לקופי האדם בני ימינו, הנתלים מהענף כלפי מטה ומתקדמים בתנופות זרוע, בעוד שעל הקרקע היא הלכה זקופה על רגליה האחוריות, אך בניגוד לבני אדם בוהן הרגל שלה הייתה פשוטה הצידה, ושימשה לשמירת שיווי משקל[32][33]. סביר שהתנועה הדו-רגלית של ארדי הייתה הרבה פחות יעילה ומהירה מההליכה והריצה של האדם, ונראה שהיא בילתה רק חלק מזמנה על הקרקע, ובשעת סכנה נמלטה מייד אל העץ הקרוב.
ממצאים מאובנים רבים מאתר הגילוי של רמידוס מראים שהוא חי בבית גידול מיוער, וסותרים השערות מקובלות מהמאה ה-20 שההזדקפות על שתיים החלה כהסתגלות לחיים בסוואנה האפריקנית. על פי השערת ה"פרנסה הזכרית" (male provisioning) של הפלאואנתרופולוג אוון לבג'וי (C. Owen Lovejoy), הליכתו הזקופה של רמידוס התפתחה כהתאמה לנשיאת מזון בידיים אל הנקבות והצעירים, במקביל להתפתחות מונוגמיה בענף האדם. גם קופי אדם מודרניים הולכים לעיתים על שתיים כאשר הם נושאים מזון בזרועותיהם, אבל הזכרים שלהם אינם מונוגמיים ואינם דואגים לצאצאיהם, פרט לגיבונים. לפי השערתו של לבג'וי, התפתחות מונוגמיה ונשיאת מזון בזכרי רמידוס אפשרה להביא מספר גדול יותר של צאצאים לבגרות, ובשל כך היה לה יתרון ברירה חזק[34][33].
האוסטרלופיתקנים והיציאה לסוואנה עריכה
- ערך מורחב – אוסטרלופיתקוס
במחצית תקופת הפליוקן, האקלים הגלובלי של כדור הארץ הפך קר ויבש יותר, וככל הנראה גרם להכחדת קופי האדם שעדיין נותרו בצפון-אסיה ובאירופה. גם באפריקה הצטמצמו שטחי היער, מה שהעניק יתרון להומינינים הולכים על שתיים, שיכלו לנצל שטחים פתוחים יותר[35], מכיוון שהליכה על שתיים חסכונית בהרבה מבחינת צריכת אנרגיה ומסייעת לצלוח הליכות ארוכות בין מקורות מים מרוחקים[36]. האוסטרלופיתקנים היו נפוצים במזרח אפריקה ודרומה במהלך הפליוקן ותחילת הפלייסטוקן, כארבעה מיליון עד מיליון שנים לפני זמננו. סביר שהם התפתחו מארדיפיתקוס רמידוס, שקדם להם רק בכמה מאות אלפי שנים, או אולי ממין דומה של ארדיפיתקוס או אורורין. האנטומיה של האוסטרלופיתקנים מוכרת היטב מהשלד המפורסם של "לוסי" ומאובנים רבים אחרים. הם הלכו זקופים על שתיים, ובניגוד לארדיפיתקוס כף הרגל שלהם כבר הייתה כמעט מודרנית בצורתה, עם קשת כף-רגל ובוהן קרובה לשאר האצבעות, כפי שניתן לראות בעקבותיהם שהתאבנו באתר לייטולי[17]. אך האוסטרלופיתקנים עדיין שמרו על יכולת מסוימת של טיפוס על עצים, כפי שניתן להסיק מזרועותיהם השוות באורכן לאורך הרגליים, אצבעותיהם הארוכות והמעוקלות, ותכונות שלד נוספות האופייניות לקופי אדם. הם חיו באזורים פתוחים יחסית, סוואנה וחורש, וקרוב לוודאי שבדומה לקופים החיים כיום בסוואנה האפריקנית הם בילו את רוב זמנם על הקרקע ומצאו שם את מזונם, אך לנו על עצים בלילה כמחסה מפני טורפים.
נפח מוחם של האוסטרלופיתקנים היה 350 עד 500 סמ"ק, כלומר עדיין כנפח מוח קוף אדם או רק במעט גדול יותר. סביר להניח שבדומה לשימפנזים בני ימינו הם השתמשו בכלים פשוטים כמו מקלות ואבנים בלתי מעובדות. ייתכן שכמה אוסטרלופיתקנים אף סיתתו אבנים באופן גס. כלי האבן המסותתים העתיקים בעולם הם בני 2.6 מיליון שנה ונמצאו באתיופיה באתר המיוחס לאוסטרלופיתקוס גרהי[37].
כיום מוכרים מרשומות המאובנים לפחות חמישה מינים של הסוג אוסטרלופיתקוס. שלושה מינים נוספים מופרדים לעיתים לסוג פראנתרופוס, שכן הם מתייחדים במבנה גוף חסון יותר עם לסתות מפותחות ושיניים טוחנות גדולות, הרומזות על התמחות בלעיסת מזון צמחי. האחרונים שבהם המשיכו להתקיים בתחילת הפלייסטוקן במקביל למיני האדם שהתפתחו מהם, אך נראה שלבסוף לא עמדו בתחרות ונכחדו לחלוטין לפני כמיליון שנה[17].
מיני אדם קדומים והגידול בנפח המוח עריכה
- ערך מורחב – אדם (סוג)
כמעט כל החוקרים כיום מסכימים שסוג האדם (Homo) התפתח מהאוסטרלופיתקנים לפני קצת יותר משני מיליון שנה, אך עדיין לא ברור היכן בדיוק ומאיזה מין[17]. המאובנים העתיקים ביותר שהתגלו מסוג האדם משויכים לרוב למינים הביליס ורודולפנסיס. הם חיו באפריקה לצד מיני אוסטרלופיתקוס ופראנתרופוס, אך הם היו בעלי מוח מעט יותר גדול, בין 510 ל-750 סמ"ק, פנים ולסת פחות בולטות, ניבים קטנים יותר, ובמבט מלמטה שורת השיניים שלהם היא בצורת קשת (פרבולה) ולא בצורה מרובעת[17]. למרות הבדלים אלו הם עדיין כה דומים לאוסטרלופיתקנים בצורת הגולגולת שלהם ובפרופורציות הגוף, עם זרועות הדומות באורכן לרגליים, עד כי מספר פלאואנתרופולוגים סבורים שיש לכלול אותם בסוג אוסטרלופיתקוס[38]. מאובני הביליס ורודולפנסיס מתגלים לרוב בסמוך לכלי אבן מסותתים מהתרבות האולדובאית ועצמות בעלי חיים שעליהן סימנים טיפוסיים של חיתוך וריסוק.
- ערך מורחב – הומו ארקטוס
הומו ארקטוס האפריקני, המסווג לעיתים כהומו ארגסטר, התפתח לראשונה במזרח אפריקה לפני כ-1.7 מיליון שנה, ככל הנראה מהביליס או רודולפנסיס, והוא המין הראשון השייך ללא עוררין לסוג האדם[17]. בדומה לאדם בן ימינו, הרגליים שלו ארוכות מן הזרועות, שלד הגוף שלו כמעט מודרני בגודלו ובצורתו, וכל ההתאמות העתיקות לטיפוס על עצים נעלמו ממנו לחלוטין. נפח המוח שלו בין 800 ו-900 סמ"ק, גדול משמעותית משל קודמיו, אך עדיין קטן משמעותית מזה של האדם המודרני. ארקטוס היה ככל הנראה המין הראשון שהשתמש באש ופיתח את התעשייה האשלית – הטכניקה הראשונה של ייצור כלי אבן שדרשה סיתות שיטתי ומוקפד של טיפוסי כלים תקניים, כמו אבן היד.
-
 הומו הביליס, גיל: 1.9 מיליון שנה, נפח מוח: 510 סמ"ק
הומו הביליס, גיל: 1.9 מיליון שנה, נפח מוח: 510 סמ"ק -
 הומו רודולפנסיס, גיל: 1.8 מיליון שנה, נפח מוח: 700 סמ"ק
הומו רודולפנסיס, גיל: 1.8 מיליון שנה, נפח מוח: 700 סמ"ק -
 הומו גאורגיקוס, גיל: 1.7 מיליון שנה, נפח מוח: 600 סמ"ק
הומו גאורגיקוס, גיל: 1.7 מיליון שנה, נפח מוח: 600 סמ"ק -
 הומו ארגסטר, גיל: 1.6 מיליון שנה, נפח מוח: 880 סמ"ק
הומו ארגסטר, גיל: 1.6 מיליון שנה, נפח מוח: 880 סמ"ק -
 הומו ארקטוס, גיל: 0.8 מיליון שנה, נפח מוח: 1,000 סמ"ק
הומו ארקטוס, גיל: 0.8 מיליון שנה, נפח מוח: 1,000 סמ"ק -
 הומו היידלברגנסיס, גיל: 0.4 מיליון שנה, נפח מוח: 1,220 סמ"ק
הומו היידלברגנסיס, גיל: 0.4 מיליון שנה, נפח מוח: 1,220 סמ"ק -
 אדם ניאנדרטלי, גיל: 0.05 מיליון שנה, נפח מוח: 1,600 סמ"ק
אדם ניאנדרטלי, גיל: 0.05 מיליון שנה, נפח מוח: 1,600 סמ"ק -
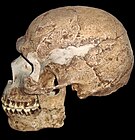 הומו ספיינס מוקדם, גיל: 0.1 מיליון שנה, נפח מוח: 1,500 סמ"ק
הומו ספיינס מוקדם, גיל: 0.1 מיליון שנה, נפח מוח: 1,500 סמ"ק





.jpg)


- ערך מורחב – הגידול בנפח המוח באבולוציה של המין האנושי
התכונה האנטומית העיקרית המגדירה את סוג האדם היא גידולו המואץ של המוח במהלך שני מיליון השנים האחרונות, החל מנפח 400 סמ"ק האופייני לאוסטרלופיתקנים ועד ממוצע של כ-1400 סמ"ק באדם המודרני. התפתחות זו מתועדת באמצעות הנפח הפנימי של גולגולות מאובנות, שרישומיהן הפנימיים אף מאפשרים להתחקות במידה מסוימת אחרי אזורי מוח ספציפיים. על אף קצבה המהיר במונחים אבולוציוניים, התפתחות זו הייתה הדרגתית מאוד, וטווח גודלי המוח של כל מין חופף חלקית את זה של הקודמים לו והמאוחרים יותר. אזור המוח שגדל יותר מכל השאר הוא קליפת המוח הגדול.

רקמות עצב צורכות אנרגיה רבה יותר מרקמות אחרות, ולכן למוח גדול יש יתרון אבולוציוני רק אם הוא מחזיר השקעה זו. מספר השערות הועלו ליתרון כזה, שקשה להכריע ביניהן וממילא אינן בהכרח סותרות:
- ההסתגלות לשימוש הולך וגובר בכלים, ובעיקר לסיתות משוכלל יותר ויותר של כלי אבן[40].
- הצורך בזיכרון טוב יותר על מנת להתמצא במרחבי הסוואנה[41][42].
- פיתוח יתירות של תאי מוח לפעולה בתנאי עקת חום (הסבר שזכה לכינוי "אני רץ, ולכן אני חושב")[43].
מיני האדם בפלייסטוקן וההגירות מאפריקה עריכה

- ערך מורחב – נדידת האדם הקדמון
עד עידן הקרח של הפלייסטוקן לפני כ-2.5 מיליון שנה, כל האבולוציה המוכרת של שושלת האדם התרחשה ביבשת אפריקה. האקלים הגלובלי של הפלייסטוקן התאפיין במספר תקופות קרות מאוד שביניהן הפוגות ממוזגות. קרחונים כיסו את מרבית האזורים הצפוניים של אירואסיה ואמריקה, ובאפריקה הפכו אזורים רבים לצחיחים יותר והקשו על תנאי החיים. מאידך ירידת מפלס פני הים במאות מטרים גרמה להצטמצמות מצרי ים, ולהיווצרות גשרי יבשה או גשרי קרחונים בין צפון-מזרח אפריקה למזרח התיכון, במצרי הבוספורוס בין אסיה לאירופה, בין דרום-מזרח אסיה לאיי אינדונזיה כמעט עד אוסטרליה, בין אירופה לאיים הבריטים, ובמצר ברינג בין סיביר לצפון אמריקה. גשרים אלו אפשרו לראשונה את התפשטותן של אוכלוסיות אדם לחלקים נרחבים של העולם[44].
שילוב עדויות המאובנים והעדויות מגנום האדם מאשר בעיקרו את השערת "היציאה מאפריקה" (Out of Africa), לפיה תולדות סוג האדם מורכבות מגלי הגירה חוזרים ונשנים מאפריקה לשאר העולם. השערה זו מוצגת לרוב כניגוד להשערת ההמשכיות האזורית (Regional Continuity), לפיה בני האדם המודרניים בכל אזור גאוגרפי התפתחו בעיקר מהמינים העתיקים יותר באותו אזור. שני גלי הגירה שמתועדים טוב יחסית הם:
- התפשטות הומו ארקטוס למזרח התיכון ולאסיה בפלייסטוקן המוקדם, כ-1.5 עד 1 מיליון שנה לפני זמננו, מתועדת על ידי ההתפשטות של מאובני ארקטוס ושל כלי אבן מסותתים, בעיקר מהתרבות האשלית, ומכונה לעיתים "היציאה מאפריקה 1" (Out of Africa I).
- התפשטות הומו ספיינס לאסיה, אוסטרליה, אירופה ואמריקה בפלייסטוקן המאוחר, כ-100 אלף עד 12 אלף שנה בלבד לפני זמננו, שיצרה את האנושות המודרנית. התפשטות זו מתועדת על ידי מאובני אדם מודרני ובעיקר באמצעות מחקר שושלות גנטיות (ראו להלן) ומכונה לעיתים "היציאה מאפריקה 2" (Out of Africa II).
בין שני גלים אלו, בפלייסטוקן התיכון, ייתכן שאירע גל נוסף של התפשטות הומו היידלברגנסיס (על-פי סיווגים אחרים ספיינס ארכאי, הומו אנטקסור או הומו רודזיינסיס) למזרח התיכון ולאירופה, שם הוא התפתח לאדם הניאנדרטלי. בנוסף, הממצאים החדשים של הומו גאורגיקוס והומו פלורסיינסיס העלו לאחרונה את ההשערה שגל הגירה עתיק מכולם אירע עוד בפלייסטוקן המוקדם מאפריקה לאסיה, של מין אדם מוקדם כלשהו כמו הביליס או רודולפנסיס, או אולי אף של אוסטרלופיתקנים[45]. נראה שכל גל הגירה כזה החליף את האוכלוסיות הקדומות יותר באותו אזור (אם היו כאלו), אך ייתכן שהתקיימו ביניהן גם שיעורים מסוימים של רביית כלאיים.
אבולוציה של המין האנושי המודרני עריכה
ספיינס ארכאי והאדם הניאנדרטלי עריכה
- ערך מורחב – הומו היידלברגנסיס
החל מתקופה של כחצי מיליון שנה לפני זמננו נמצאו באפריקה, אירופה ואסיה מאובני אדם רבים שנפח המוח שלהם כ-1,200 סמ"ק – גדול מהטווח התחתון של מוחות בני אדם נורמלים כיום – ולעיתים רחוקות אף מעל ל-1,350 סמ"ק, שהוא הממוצע המודרני. חלק ממאובנים אלו מסווגים על ידי חלק מהחוקרים תחת המונח החצי-רשמי "הומו ספיינס ארכאי". השימוש בשם המין "ספיינס" פירושו שלדעת החוקרים, בני אדם אלו כבר היו יכולים להעמיד ללא קושי צאצאים פוריים עם בני אדם מודרניים אילו חיו היום. המונח "ארכאי" פירושו שהם עדיין נבדלים באנטומיה שלהם מבני אדם מודרניים, למשל הם חסרים עדיין את בליטת הסנטר על הלסת התחתונה ורכסי הגבות שלהם מפותחים מאוד. תכונות אלו טיפוסיות לארקטוס, היידלברגנסיס והאדם הניאנדרטלי, ואכן רוב מאובני "ספיינס ארכאי" מסווגים על ידי חוקרים אחרים כארקטוס, כהיידלברגנסיס או כ"קדם ניאנדרטלים", וההבדל בין טיפוסים אלו הוא כה הדרגתי שקשה מאוד להגדירו[46]. בדומה לארקטוס, מאובנים אלו עדיין מקושרים לרוב עם תרבות כלי האבן האשלית.
- ערך מורחב – האדם הניאנדרטלי
החל מתקופה של כ-250 אלף שנה לפני זמננו, בפלייסטוקן המאוחר, מאובני ספיינס ארכאי במזרח התיכון ובאירופה מראים השתנות הדרגתית לאוכלוסייה המוכרת כיום כאדם הניאנדרטלי, שבצורתה המובהקת התקיימה כ-120 אלף עד 20 אלף שנה לפני זמננו. לכן חוקרים "אחדנים" מסווגים את הניאנדרטלים כתת-מין של ספיינס, מכנים אותם "הומו ספיינס ניאנדרתלנסיס" ומשייכים אותם לקבוצת ספיינס הארכאי. לעומת זאת חוקרים "פצלנים" סבורים שבכל זאת יש לסווגם כמין נפרד: "הומו ניאנדרתלנסיס". חקר הגנום הניאנדרטלי בשנים האחרונות מראה שהניאנדרטלים היו אוכלוסייה נפרדת מאוד מהאדם המודרני, ועם זאת, המחקר העדכני ביותר בנושא מעריך שכ-1%–4% מהשונות הגנטית של בני-אדם החיים כיום מחוץ לאפריקה ניתן לייחס לאללים ניאנדרטלים, ומכאן ששיעור נמוך של רביית כלאיים פורייה התקיים בין הניאנדרטלים לספיינס המודרני בעת התפשטותו של האחרון מאפריקה.
הניאנדרטלים היו בעלי נפח מוח דומה לזה של בני אדם בני ימינו, ואפילו גדול ממנו במקצת, בממוצע, אם כי יתרון זה נבע מגופם הגדול יותר בממוצע של הניאנדרטלים ולא ממוח גדול יותר באופן יחסי. גם תרבות כלי האבן האופיינית לניאנדרטלים – התרבות המוסטרית המשויכת לתקופה הפלאוליתית התיכונה – מתקדמת מהתרבות האשלית המשויכת לתקופה הפלאוליתית התחתונה. עם זאת, חפצי אמנות, קישוטים וגילויים אחרים של חשיבה סימבולית עדיין נדירים ביותר בתרבות המוסטרית.

הופעת המין האנושי המודרני עריכה
- ערך מורחב – אדם
טיפוסי ספיינס ארכאי שחיו במזרח התיכון ובאירופה התפתחו שם לניאנדרטלים, ובמקביל, טיפוסי ספיינס ארכאי שחיו באפריקה התפתחו שם לבני אדם מודרניים. ההתפתחות באפריקה מוכרת פחות עקב המיעוט היחסי של חפירות פלאואנתרופולוגיות וארכאולוגיות שם. מאובנים שהתגלו באתיופיה על ידי הצוות של טים וייט באזור החפירה הקבוע שלו באוואש התיכון, הם בעלי מוח גדול (כ-1,450 סמ"ק) אבל מאפייני שלד ארכאיים, וסווגו כתת-מין הומו ספיאנס אידלטו. הם מתוארכים ל-160 אלף שנה לפני זמננו, התארוך הידוע המאוחר ביותר של בני אדם שאינם מודרניים לחלוטין באפריקה[47].


"מודרניות" של בני אדם ניתן להגדיר לפחות בשלושה מובנים שונים: במובן האנטומי על פי תצורת השלד שלהם, במובן הגנטי על פי רצף הגנום שלהם, ובמובן ההתנהגותי על פי התרבות שלהם. מאובני אדם הזהים אנטומית לבני אדם בני ימינו מוגדרים במינוח הטקסונומי כתת-מין "הומו ספיינס ספיינס" (Homo sapiens sapiens). המאובנים העתיקים ביותר שסווגו כך התגלו באתר קיביש סמוך לנהר אומו בדרום-אתיופיה, עוד בשנות ה-60 של המאה ה-20 על ידי משלחות של ריצ'רד ליקי, אולם רק לאחרונה תארוך רדיומטרי מתקדם בשיטת ארגון-ארגון העריך את גילם ב-190 אלף שנה[48]. בהשוואה לניאנדרטלים וספיינס ארכאי, התכונות האנטומיות המציינות את האדם המודרני הן:
- גולגולת גבוהה יותר (בממד האנכי) וקצרה יותר (מפנים לאחור).
- רכסי הגבות "נבלעים" לחלוטין או כמעט לחלוטין על ידי מצח גבוה ובולט.
- הלסתות קטנות עד כדי כך שהשיניים הטוחנות האחוריות ביותר (שיני בינה) בוקעות בגיל מאוחר יחסית, ולפעמים אינן בוקעות כלל.
- בליטת סנטר מפותחת בלסת התחתונה.
- מבנה שלד עדין יותר באופן כללי, עם עצמות דקות יותר ורכסי עצם (המשמשים כמאחז לשרירים) פחות מפותחים.
מחקרים גנטיים באוכלוסיות אדם בנות ימינו, המסתמכים על רצפי דנ"א מיטוכונדרי, העובר בירושה דרך השושלת הנקבית בלבד, רצפי כרומוזום Y, העובר בירושה דרך השושלת הזכרית בלבד, ולאחרונה גם על רצפי דנ"א רגיל, מראים כולם באופן בלתי תלוי כי ה"שורש" של כל השושלות האנושיות דומה ביותר לרצפים של בני אדם אפריקנים. ההערכה המעודכנת לגילה של חוה המיטוכונדרית, אם משותפת של כל בני האדם החיים היום, היא כ-200 אלף שנה. גילו של אדם כרומוזום Y, האב משותף של כל בני האדם החיים היום, הוא לפי הערכות עדכניות כ-142,000 שנה. מכאן שיש כיום הסכמה בין עדויות המאובנים לעדויות הגנומיות בנוגע למוצאה האפריקני של האנושות המודרנית. מעקב אחר השושלות הגנטיות מאפשר אף לתארך את הגירת האדם המודרני מאפריקה לשאר היבשות, ומועדי ההגירה למזרח התיכון (לפני כ-100 אלף שנה), לאוסטרליה (לפני כ-50 אלף שנה), לאירופה (לפני כ-35 אלף שנה) וליבשת אמריקה (לפני כ-15 אלף שנה) אף הם תואמים ברובם את תארוך הופעתם הראשונה של מאובני אדם מודרני אנטומית במקומות אלו. שרידי האוכלוסייה האירופית הם הראשונים שהתגלו עוד במאה ה-19, ועד היום מכנים אותם לעיתים "קרו-מניון" על שם מערה בצרפת שבה נמצאו.
- ערך מורחב – אמנות פרהיסטורית
עם זאת, מודרניות גנטית ואנטומית אין פירושה בהכרח גם מודרניות התנהגותית[49]. בארץ ישראל בעיקר נמצאו מהתקופה שבין 100 אלף ל-50 אלף שנה לפני זמננו מאובני ניאנדרטלים וגם מאובני אדם מודרני אנטומית, שחיו באותו זמן ואולי אף זה לצד זה, ועדיין כולם עם שרידים ארכאולוגים אופייניים לאותה תרבות – התרבות המוסטרית. רק בממצאי התקופה הפלאוליתית העליונה נפוצות כל אותן עדויות למודרניות תרבותית שאופייניות גם לאוכלוסיות ציידים-לקטים בנות זמננו: להבי אבן עדינים בעלי סיתות משוכלל ביותר, כלי עצם, קישוטים וחפצי אמנות, כלי נגינה, סמלים גאומטריים, חומרי צבע כמו אוכרה, קבורה טקסית וגילויים אחרים של חשיבה סימבולית. הממצאים העתיקים המעידים ללא ספק על מודרניות התנהגותית הם חרוזי צדף וממצאים אחרים ממערת הגדי (סח'ול) בנחל מערות בארץ-ישראל, מלפני 130 אלף עד 100 אלף שנים[50], וממערת בלומבוס בדרום אפריקה, מלפני 85 אלף עד 75 אלף שנים. ואולם במזרח התיכון, אירופה ואסיה נראה כי התקופה הפלאוליתית העליונה החלה רק לפני 60 אלף עד 40 אלף שנה. יש המשערים שהיא מעידה על הופעתה של שפה סימבולית מודרנית[51].
הרכב גנטי של אוכלוסיות אדם בנות ימינו עריכה
- ערך מורחב – גזע (אדם)
רצף הגנום של אוכלוסיות אדם מודרניות משקף את האבולוציה שלהן ואת מיקומן הגאוגרפי. זאת משום שאנשים נוטים יותר להעמיד צאצאים עם אנשים שגרים סמוך אליהם, כך שבמשך הדורות נוצר מתאם בין אזור המוצא לרצף הגנום[52]. מחקרים מהשנים האחרונות שמודדים מספר גדול מאוד (כמה אלפים לפחות) של סמנים גנטיים מסוגלים לעיתים קרובות לזהות את אזור מוצאם של נבדקים שאינם בני תערובת. עם זאת, מרבית הגנטיקאים כיום אינם משתמשים עוד במונח "גזע" לתיאור מתאם זה, משום שהמתאם הוא רציף ואיננו מתחלק באופן בדיד לקטגוריות מסורתיות כמו שחורים ולבנים[53][52]. המינוח המסורתי של גזעים גם איננו מקביל בהכרח לתת-מינים המשמשים במינוח הזואולוגי בשביל מיני בעלי חיים אחרים. זאת משום שהאנושות המודרנית צעירה בהשוואה למרבית המינים, כך שהשונות הגנטית בה נמוכה יחסית למרות תפוצתה הכלל-עולמית[52]. גם הגירות ונדידות עמים תכופות לאורך הפרהיסטוריה וההיסטוריה, המלוות לרוב בנישואי תערובת, תרמו להקטנת ההבדלים הגנטיים בין אזורים גאוגרפיים ואוכלוסיות אתניות. לשם השוואה, מחקרים גנטיים במין השימפנזה המצוי מצאו שמספר תת-מינים שלו באפריקה לבדה התפצלו לפני מאה אלף עד מיליון שנים[54][55], בעוד שזמני ההתפצלות בין "גזעי" אדם על פני כדור הארץ כולו נמדדים באותן שיטות בעשרות אלפי שנים לכל היותר.
אבולוציה של האדם בימינו עריכה
סברה שגויה הרווחת בציבור היא שאבולוציה איננה מתרחשת עוד בקרב בני אדם כיום, אם משום שאין רואים אותה בפעולה ואם משום שחידושי הטכנולוגיה, על פי סברה זו, מבטלים את הברירה הטבעית. אך למעשה, כל שינוי בשכיחויות אללים באוכלוסייה מוגדר כאבולוציה, ושינויים כאלו אכן מתרחשים, אלא שהם אורכים אלפי שנים לכל הפחות, כך שקשה להבחין בהם. גנטיקאים העלו את ההשערה שקצב האבולוציה האנושית אף הואץ בעשרות אלפי השנים האחרונות, כתוצאה מהגידול במספר הנפשות שגורם לגידול במספר המוטציות החדשות המופיעות בכלל האוכלוסייה[56].
- ערך מורחב – רגישות ללקטוז
דוגמה נחקרת היטב לאבולוציה מודרנית היא התפתחות הסבילות לסוכר הלקטוז במבוגרים, המאפשרת להם לעכל חלב של בהמות שונות (במצב הטבעי רק תינוקות מסוגלים לעכל את סוכר החלב). מחקר גנטי הראה שאבולוציה זו היא תוצאה של מוטציות בגורם שעתוק המדכא את ייצור אנזים הלקטאז, מוטציות שהופיעו לראשונה לפני כמה אלפי שנים, בראשיתה של המהפכה החקלאית. בתרבויות אנושיות המגדלות בהמות חלב, מוטציות אלו כבר התפשטו למרבית האוכלוסייה כתוצאה מברירה טבעית, אך בתרבויות שאינן צורכות מוצרי חלב אין למוטציות אלו יתרון כלשהו, ולכן הן נדירות[57][58]. במחקר פלאוגנטי של 230 גנומים משלדי בני-אדם עתיקים באירואסיה נמצאו סימנים רבים לפעולתה של ברירה טבעית, החזק שבהם במוטציה הגורמת סבילות ללקטוז[59].
מחלוקות, כשלים ותפיסות מוטעות עריכה
ההתנגדות לרעיון האבולוציה של האדם עריכה
- ערך מורחב – ההתנגדות לתורת האבולוציה
רעיונות האבולוציה, ובמיוחד משמעותם לגבי האבולוציה של האדם, עוררו מאז פרסומם ועדיין מעוררים פולמוס חריף. על עצם התפתחותם של בני אדם מקופי אדם קדומים יש כיום קונצנזוס מדעי מוחלט[60]. מזה עשרות רבות של שנים לא פורסם מאמר מדעי כלשהו שעבר ביקורת עמיתים שחולק על כך, או אף מטיל בכך ספק משמעותי כלשהו. עם זאת, פרטים רבים בהתפתחות זו אינם ידועים, אינם מובנים כל צורכם או שנויים במחלוקת.
גם מרבית המדענים הדתיים בתחומים הרלוונטיים אינם מערערים כיום על האבולוציה של האדם. המדענים הדתיים בארצות הברית המצהירים כי אין כיום מקום לספק בדבר מוצאו של האדם מקופי אדם קדומים הם למשל פרנסיס קולינס, גנטיקאי נוצרי אוונגליסטי שעמד בראש פרויקט גנום האדם הבינלאומי וכיום עומד בראש המכונים הלאומיים לבריאות[א]; קן מילר, ביולוג קתולי המוביל מאבק ציבורי נגד בריאתנות[ב]; ומייקל ביהי, ביוכימאי קתולי הידוע בתמיכתו בתכנון תבוני[ג]. גם בישראל, רוב המדענים הדתיים נוקטים בגישות שונות ליישוב אמונתם עם האבולוציה[64], וכן הרב דניאל הרשקוביץ, פרופסור למתמטיקה בטכניון שהיה שר המדע, הביע את דעתו כי אין לשלול שמוצא האדם הוא מקופי אדם, ואף הציע מקורות ביהדות התומכים בכך[65].
למרות זאת ציבורים נרחבים בעולם, בעיקר מפלגים שמרניים של הדתות המונותאיסטיות, עדיין דוחים לחלוטין את רעיון האבולוציה של האדם. בסקר שנערך בשנת 2005, במרבית מדינות מערב אירופה למעלה מ-60% מהמבוגרים קיבלו כאמת את ההצהרה "בני אדם התפתחו ממינים קודמים של בעלי חיים", אך בארצות הברית רק כ-40% ובטורקיה רק כ-25%[66]. סקר זה לא נערך בישראל, אך בסקר אחר של מרכז המחקר פיו שנערך בישראל בשנת 2016 הסכימו 53% מהיהודים ו-37% מהערבים כי "בני אדם ויצורים חיים אחרים התפתחו באבולוציה במשך הזמן"[67]. בארצות הברית המתנגדים הם בעיקר נוצרים מהזרמים האוונגליסטיים, בעוד האפיפיור מצהיר כי אין סתירה בין הנצרות הקתולית והאבולוציה של האדם. גם בציבור הדתי והמסורתי בישראל רבים דוחים את רעיון האבולוציה של האדם[ד], ולעומתם אחרים כמו הרב אורי שרקי סבורים שאין כל מניעה שהגוף הפיזי שבו נפח הבורא רוח חיים היה קוף או אדם קדמון.
בישראל, האבולוציה של האדם אינה נכללת בתוכנית לימודי החובה של משרד החינוך, אף כי אין מדיניות רשמית נגד מורים המלמדים אותה ביוזמתם. בשנת 2014 פורסם שקיימת תוכנית במשרד החינוך לכלול אבולוציה בלימודי מדעים בחטיבות הביניים, אך נושא האבולוציה של האדם לא נזכר בה. חבר ועדת משרד החינוך למקצוע המדעים חגי נצר צוטט כי "אין התייחסות ישירה לנושא הזה מפני שמדובר בנושא רגיש מאוד במדינת ישראל"[69]. מיעוט זעיר של תלמידי תיכון בישראל לומדים אבולוציה של האדם כחלק מפרק רשות על אבולוציה במסגרת ההקבצה הגבוהה ביותר של בחינות הבגרות בביולוגיה.
רבים בתחומי המדע והדת כאחת אינם שוללים את רעיון האבולוציה של בני אדם, אך סוברים שאין די בו על מנת להסביר את "מותר האדם", היינו ייחודו הנפשי או המוסרי של האדם. תורת האבולוציה אינה מתיימרת לספק הסבר מלא ומוחלט לכל תכונותיו של האדם.

תפיסת האבולוציה כ"סולם" וכ"שיח" עריכה
בציבור הרחב עדיין נפוצה תפיסה שגויה שרווחה בקהילה המדעית בתחילת המאה ה-20, לפיה האבולוציה מתנהלת באופן סדרתי מצורות פשוטות יותר למורכבות יותר, התפיסה שסטיבן גולד כינה "הסולם"[70]. בהקשר של האבולוציה של האדם, תפיסה פשטנית זו מתבטאת בתמונת "מצעד הקדמה", אחד הסמלים הגרפיים המוכרים ביותר בתרבות המערבית, שרבים בציבור נוטים לזהותו כמעט אוטומטית עם רעיון האבולוציה. במציאות, כל הקבוצות המוכרות היטב מרשומות המאובנים, כגון סוסיים או דינוזאורים, מפגינות אבולוציה מסועפת מאוד שאיננה בהכרח "מתקדמת" ליצורים מורכבים יותר או מותאמים יותר לסביבתם – התפיסה שגולד כינה "השיח". האבולוציה של האדם אינה יוצאת דופן מבחינה זו: היא מאוד מסועפת ורבים מהמינים הכלולים בה התקיימו במקביל במשך זמן רב[71]. מי שאינו מבין עניין זה עלול להסיק, למשל, שהכחדתו של האדם הניאנדרטלי מעידה בהכרח על כך שהוא היה פחות מפותח מהאדם המודרני, או שהוא היה אביו הקדמון של האדם המודרני, אך מסקנות מעין אלו נחשבות כיום שגויות.
"החוליה החסרה" עריכה

בסוף המאה ה-19 ובתחילת המאה ה-20 התנהלו ויכוחים סוערים בתחום הפלאואנתרופולוגיה מיהו המאובן שזכאי לתואר "החוליה החסרה" בין קוף האדם לאדם. בין הממצאים שטענו בשעתם לתואר זה היו האדם הניאנדרטלי, פיתקאנתרופוס ארקטוס (כיום מסווג כהומו ארקטוס מג'אווה), אדם פקין (כיום מסווג כהומו ארקטוס מסין), איש פילטדאון (שהסתבר כזיוף) ואוסטרלופיתקוס אפריקנוס (מאובן ה"ילד מטאונג" שגילה ריימונד דארט). בוויכוחים אלו היו מעורבים ככל הנראה גם שיקולים בלתי מדעיים, למשל מדענים בריטים נטו לתמוך באיש פילטדאון הבריטי, בעוד מדענים גרמנים נטו לתמוך באדם הניאנדרטלי שהתגלה בגרמניה, והאירופים בכללם נטו להתעלם ממועמדים אסייתיים ואפריקניים. מושג החוליה החסרה גם זכה לחשיפה רבה בתקשורת, ונעשה בו שימוש מוגזם ומוטעה על ידי עיתונאים ושרלטנים, שלעיתים אף המציאו "חוליות חסרות" משל עצמם, למשל במקרה של "איש נברסקה".
בספרות המדעית פסק לחלוטין השימוש במונח "החוליה החסרה" בעשרות השנים האחרונות. סיבה אחת לכך היא ריבוי מיני ההומינידים המאובנים שהתגלו. משמעות הריבוי היא שלא מין אחד אלא מספר מינים (על פי מרבית הסיווגים וההשערות לפחות חמישה וקרוב לוודאי יותר) מהווים מאובני מעבר בין האדם המודרני לבין האב הקדמון המשותף לאדם ולשימפנזה, וממילא לא קיים מין יחידי שניתן לכנותו "החוליה החסרה" בה"א הידיעה. סיבה נוספת היא המבנה המסועף ("דמוי שיח" ולא "דמוי סולם") של האבולוציה האנושית, כך שכמעט בלתי אפשרי לקבוע בוודאות איזה מאובן נמצא בדיוק על השושלת המפותלת המובילה לאדם המודרני ואיזה מאובן נמצא על הסתעפות צדדית מעט. למרות זאת, בתקשורת ובציבור עדיין רווח שימוש מוטעה במונח "החוליה החסרה", בעיקר ב"סקופים" עיתונאיים המדווחים באופן שגוי על גילוי מאובנים חדשים של הומינידים. בעיני הקהילה המדעית בתחום, מי שעושה שימוש במונח לרוב מתייג את עצמו כחובבן במקרה הטוב וכשרלטן במקרה הגרוע.
"האם מוצא האדם הוא מן הקוף?" עריכה
שאלה זו, הנפוצה בציבור הרחב, יוצאת לפעמים מנקודת הנחה שגויה שעל פי תורת האבולוציה המודרנית מוצא האדם הוא ממין קוף הקיים כיום. ניסוח יותר מדויק יהיה "מוצא האדם המודרני וקופי האדם המודרניים הוא מאב קדמון משותף". עם זאת, יש לזכור שאותו אב קדמון היה אף הוא קוף אדם, אם כי לא ממין החי כיום. לכן מדויק גם לומר כי "מוצא האדם הוא ממיני קופים קדומים".
יתר על כן, במובן הזואולוגי אף נכון לומר שהאדם הוא קוף בעצמו. מבחינה טקסונומית האדם שייך בין השאר לתת-סדרה Simiiformes (פירוש מילולי בלטינית: "בעלי צורת קוף"), בתוכה לעל-משפחה קופי אדם (בלטינית Hominoidea שפירושו המילולי "דמויי אדם") ובתוכה למשפחת ההומינידים (הכוללת גם את השימפנזה, הגורילה והאורנגאוטן). באותו מובן נכון גם לומר שהאדם הוא פרימט, יונק, חולייתן, בעל-חיים, וכדומה.
ראו גם עריכה
לקריאה נוספת עריכה
- חוה יבלונקה (עורכת), אבולוציה, יחידה 7 - אבולוציה של האדם, מוציא לאור: האוניברסיטה הפתוחה, תשנ"ז 1997. ISBN 965-06-0372-7. (קריאת הספר בתצוגה מקדימה באתר "גוגל ספרים"
 )
) - בריאן סייקס, שבע בנות חוה, עם עובד, 2001
- פראנס דה-ואל, הקוף שבתוכנו, הוצאת כתר, 2009. (דף הספר באתר "גוגל ספרים" )
- אברהם רונן, "חשיבותה של ארץ ישראל בתולדות אדם הקדמון", קדמוניות, 144, 2012, עמ' 65–70
- Bernard Wood (2010) Reconstructing human evolution: Achievements, challenges, and opportunities, PNAS, Vol. 107, pp. 8902–8909
- Morris Goodman and Kirstin N. Sterner (2010) Phylogenomic evidence of adaptive evolution in the ancestry of humans, PNAS, Vol. 107, pp. 8918–8923
- Encyclopedia of Human Evolution and Prehistory, by Eric Delson, Ian Tattersall, John A. Van Couvering (Eds), 2nd Edition, 2006. Garland Publishing, NY
- Ralph L. Holloway et al. (2008) Evolution of the Brain, in Humans – Paleoneurology, In: Binder MD, Hirokawa N, Windhorst U, Hirsch MC, editors. Encyclopedia of Neuroscience. Berlin: Springer-Verlag
- Ian Tattersall (2008) An Evolutionary Framework for the Acquisition of Symbolic Cognition by Homo sapiens, Comparative Cognition and Behavior Reviews, Volume 3, pp. 99–114
קישורים חיצוניים עריכה
- סיינטיפיק אמריקן, אחוז אחד של אנושיות, באתר ynet, 28 באוגוסט 2009
- אבולוציה של האדם באתר מוזיאון הסמיתסוניאן (באנגלית)
- אבולוציה של האדם באתר המוזיאון האוסטרלי (באנגלית)
- הניצוץ האנושי - תוכניות וידאו בהנחיית אלן אלדה על אבולוציה של האדם ומדעי המוח, אתר רשת הטלוויזיה PBS האמריקנית (באנגלית)
- אתר המכון למוצא האדם בהשתתפות דונלד ג'והנסון באוניברסיטת המדינה של אריזונה (באנגלית)
- בלוג על פלאואנתרופולוגיה, גנטיקה ואבולוציה של האדם של הפלאואנתרופולוג ג'ון הוקס (באנגלית)
- אתר אנתרופולוגיה. נט (באנגלית)
- האבולוציה - עליית האדם, באתר "הידען"
- קייט וונג, הקומזיץ הראשון בהיסטוריה, במדור סיינטיפיק אמריקן של מכון דוידסון, 1 בדצמבר 2013
- אבולוציה של האדם באתר מדע גדול, בקטנה
- נתיב המאובנים: העדות לאבולוציית האדם, תערוכה, בית האקדמיה הלאומית הישראלית למדעים, ירושלים, נפתח 3 פברואר 2019
- מאיפה באנו? המדריך המלא לאבולוציה האנושית | הדרך שעבר האדם ב-4.5 מיליון שנים | ההוכחות שדרווין צדק הולכות ונערמות | התשובות המדעיות לכל הטיעונים שמפקפקים באבולוציה | אבולוציה נהפכה למלה גסה בישראל | במלחמתה באבולוציה, הדת תצטרך להסתגל או להיכחד | 12 שנות לימוד בלי מילה על אבולוציה | ההתקפה על האבולוציה היא התקפה על הלא־מודע | למה אנחנו מפחדים מדרווין ולמה אנחנו לא יכולים לברוח ממנו? | באתר הארץ, 2 בספטמבר 2020
- אבולוציה של האדם, באתר אנציקלופדיה בריטניקה (באנגלית)
 אבולוציה של אדם, דף שער בספרייה הלאומית
אבולוציה של אדם, דף שער בספרייה הלאומית
ביאורים עריכה
- ^ בספרו "שפתו של אלוהים" קולינס כותב בין השאר כי "חקר הגנום מוליך באופן בלתי-נמנע למסקנה שאנו בני האדם חולקים אבות משותפים עם שאר היצורים החיים".[61]
- ^ קן מילר ציטט את השוואת הגנום של קופי האדם והאדם כעדות ה"מאשרת באופן ספקטקולרי" את מוצאם המשותף, בעדותו כעד מומחה במשפט הפדרלי קיצמילר-נגד-דובר 2005 בארצות הברית.[62]
- ^ ביהי כתב: "לדוגמה, בני-אדם ושימפנזים חולקים עותק שבור של הגן אשר ביונקים אחרים משתתף בייצור ויטמין סי... קשה לדמיין כיצד תיתכן ראייה חותכת יותר למוצא המשותף של שימפנזים ובני-אדם... על-אף כמה שאלות פתוחות שנותרו, אין סיבה לפקפק שדרווין צדק בנקודה זו, וכל היצורים עלי אדמות הם קרובי משפחה ביולוגיים".[63]
- ^ בסקר של מרכז המחקר פיו ב-2016, 96% מן החרדים, 85% מן הדתיים, 58% מן המסורתיים ו-14% מן החילונים ענו כי "בני אדם ויצורים חיים אחרים התקיימו בצורתם הנוכחית מאז ראשית הזמן".[68]
הערות שוליים עריכה
- ^ יואל רק, על אנשים ועל מאובנים, אוניברסיטת תל אביב, דקות 34:03-28:12 + דקה 61 ואילך
- ^ 1 2 The Chimpanzee Sequencing and Analysis Consortium (2005) "Initial sequence of the chimpanzee genome and comparison with the human genome" Nature. Vol. 437, pp. 69-87
- ^ 1 2 3 Carroll, S. B. (2003) "Genetics and the making of Homo sapiens", Nature. Vol. 422, pp. 849-857. DOI: 10.1038/nature01495
- ^ ד"ר יונת אשחר, הגודל לא תמיד קובע, במדור "חדשות מדע" באתר של מכון דוידסון לחינוך מדעי, 12 במאי 2021
- ^ 1 2 3 Delson, E. and Tattersall, I.(2000) “Primates”; in the Encyclopedia of Human Evolution and Prehistory, 2nd Ed (Eds: Delson, E. Tattersal, I. Van Couvering J. A. & Brooks, A. S.). Garland Publishing, NY. pp 1219-1228.
- ^ "מחקר חשף את השינוי הגנטי שגרם לכך שלבני אדם אין זנב". הארץ. נבדק ב-2024-03-01.
- ^ Gallup, G. G. (1970) "Chimpanzees: Self-Recognition" Science. Vol. 167, pp. 86-87; doi: 10.1126/science.167.3914.86
- ^ Call, J. and Tomasello, M. (2006) "Does the chimpanzee have a theory of mind? 30 years later.", Trends in Cognition Science. Vol. 12, pp. 187-192
- ^ Schmelz, M. et al. (2011) "Chimpanzees know that others make inferences", Proceedings of the National Academy USA. Vol. 108 (7), pp. 3077-3079
- ^ Haslam, M. et al. (2009) "Primate archaeology" Nature Reviews, Vol. 460, pp 339-344, doi:10.1038/nature08188
- ^ Pruetz, D. J. and Bertolani, P. (2007) "Savanna Chimpanzees, Pan troglodytes verus, Hunt with Tools", Current Biology. Vol. 17, pp. 412-417; doi: 10.1016/j.cub.2006.12.042
- ^ Horner, V. and de Waal, F. B. (2009) "Controlled studies of chimpanzee cultural transmission", Progress in Brain Research. Vol. 178, pp. 3-15
- ^ Toth, N. et al. (1993) "Pan the Tool-Maker: Investigations into the Stone Tool-Making and Tool-Using Capabilities of a Bonobo (Pan paniscus)", Journal of Archaeological Science. Vol. 20, pp. 81-91; doi:10.1006/jasc.1993.1006
- ^ E. Sue Savage-Rumbaugh et al. (2001) Apes, Language, and the Human Mind, Oxford University Press.
- ^ Rumbaugh, D. M. et al. (1987) "Summation in the chimpanzee (Pan troglodytes)", Journal of Experimental Psychology: Animal Behavior Processes. Vol. 13, pp. 107-115; doi: 10.1037/0097-7403.13.2.107
- ^ Biro, D. and Matsuzawa, T. (2007) "Use of numerical symbols by the chimpanzee ( Pan troglodytes ): Cardinals, ordinals, and the introduction of zero", Animal Cognition. Vol. 4, pp. 193-199; doi: 10.1007/s100710100086
- ^ 1 2 3 4 5 6 7 Wood, B. (2010) "Reconstructing human evolution: Achievements, challenges, and opportunities", Proceedings of the National Academy of Sciences. Vol. 107, pp. 8902-8909
- ^ Patterson, N. et al (2006) "Genetic evidence for complex speciation of humans and chimpanzees", Nature . Vol. 441, pp. 1103-1108; doi:10.1038/nature04789
- ^ Yang, Z. (2002) "Likelihood and Bayes Estimation of Ancestral Population Sizes in Hominoids Using Data From Multiple Loci", Genetics. Vol. 162, pp. 1811-1823
- ^ 1 2 Cyran, K. A. and Kimmel, M. (2010) "Alternatives to the Wright-Fisher model: the robustness of mitochondrial Eve dating", Theoretical population biology. Vol. 78(3), pp. 165-172
- ^ Krause, J. et al (2010) "The complete mitochondrial DNA genome of an unknown hominin from southern Siberia", Nature . Vol. 464, pp. 894-897; doi:10.1038/nature08976
- ^ Ohta, Y. and Nishikimi, M. (1999) "Random nucleotide substitutions in primate nonfunctional gene for L-gulono-gamma-lactone oxidase, the missing enzyme in L-ascorbic acid biosynthesis", Biochimica et Biophysica Acta. Vol. 1472, pp. 408-411
- ^ Chang, L. Y, and Slightom, J. L. (1984) "Isolation and nucleotide sequence analysis of the beta-type globin pseudogene from human, gorilla and chimpanzee.", Journal of Molecular Biology. Vol. 180(4), pp. 767-784
- ^ Pollard, K. S. et al. (2006) "Forces Shaping the Fastest Evolving Regions in the Human Genome", PLoS Genetics. Vol. 2(10), e168
- ^ Pollard, K. S/ et al. (2006) "An RNA gene expressed during cortical development evolved rapidly in humans", Nature . Vol. 443, pp. 167-172; doi:10.1038/nature05113
- ^ Enard, W. et al. (2009) "A Humanized Version of Foxp2 Affects Cortico-Basal Ganglia Circuits in Mice" Cell. Vol. 137(5), pp. 961-971; doi:10.1016/j.cell.2009.03.041
- ^ Somel, M. et al. (2009) "Transcriptional neoteny in the human brain"Proceedings of the National Academy of Sciences. Vol. 106 (14), pp. 5743-5748. DOI: 10.1073/pnas.0900544106
- ^ Kunimatsu, Y. (2007) "A new Late Miocene great ape from Kenya and its implications for the origins of African great apes and humans", Proceedings of the National Academy of Sciences. Vol. 104(49), pp. 19220-19225
- ^ Brunet, M. et al. (2002) "A new hominid from the Upper Miocene of Chad, Central Africa" Nature. Vol. 418, pp. 145-151; doi: 10.1038/nature00879
- ^ Senut, B. et al. (2002) "First hominid from the Miocene (Lukeino Formation, Kenya)", Comptes Rendus de l’Académie des Sciences - Series IIA - Earth and Planetary Science. Vol. 332 (2), pp. 137-144; doi: 10.1016/S1251-8050(01)01529-4
- ^ Haile-Selassie, Y. (2001) "Late Miocene hominids from the Middle Awash, Ethiopia", Nature. Vol. 412, pp. 178-181; doi: 10.1038/35084063
- ^ 1 2 3 White, D. T. et al. (2009) "Ardipithecus ramidus and the Paleobiology of Early Hominids" Science. Vol. 326 (5949), pp. 75-86; doi: 10.1126/science.1175802
- ^ 1 2 Lovejoy, C. O. (2009) "Reexamining Human Origins in Light of Ardipithecus ramidus", Science. Vol. 326 (5949), pp. 74e1-74e8; doi: 10.1126/science.1175834
- ^ Lovejoy, C. O. (1981) "The Origin of Man", Science. Vol. 221, pp. 341 - 350; doi: 10.1126/science.211.4480.341
- ^ Van Couvering, J. A. (2000) “Pliocene”; in the Encyclopedia of Human Evolution and Prehistory, 2nd Ed (Eds: Delson, E. Tattersal, I. Van Couvering J. A. & Brooks, A. S.). Garland Publishing, NY. pp 1185-1188.
- ^ יואל רק, על אנשים ועל מאובנים- הרצאה ראשונה בסדרה, אוניברסיטת תל אביב, במיוחד בדקות 59–65
- ^ Semaw, S. (2000) "The World's Oldest Stone Artefacts from Gona, Ethiopia: Their Implications for Understanding Stone Technology and Patterns of Human Evolution Between 2.6–1.5 Million Years Ago ", Journal of Archaeological Science. Vol. 27(12), pp. 1197-1214; doi:10.1006/jasc.1999.0592
- ^ Wood, B. and Collard, M. (1999) "The Human Genus", Science. Vol. 284(5411), pp. 65-71; doi: 10.1126/science.284.5411.65
- ^ Schoenemann, P. T (2006) "Evolution of the Size and Functional Areas of the Human Brain", Annual Reviews in Anthropology. Vol. 35, pp. 379-406; doi:10.1146/annurev.anthro.35.081705.123210
- ^ Stout, D. et al. (2008) "Neural correlates of Early Stone Age toolmaking: technology, language and cognition in human evolution", Philosophical Transactions of the Royal Society B. Vol. 363 (1499), pp. 1939-1949; doi:10.1098/rstb.2008.0001
- ^ G. S. Krantz (1968) Brain size and hunting ability in earliest man. Current Anthropology 9:450-451
- ^ Eckhardt, R. B. (1987) Was plio-pleistocene hominid brain a pleiotrpic effect of adaptation to heat stress? Anthropology 45:193-201
- ^ Fialkowski, K. R. (1978) Early hominid brain evolution and stress, Studies in Physical Anthropology 4:87-92
- ^ Van Couvering, J. A. (2000) “Pleistocene”; in the Encyclopedia of Human Evolution and Prehistory, 2nd Ed (Eds: Delson, E. Tattersal, I. Van Couvering J. A. & Brooks, A. S.). Garland Publishing, NY. pp 1168-1177.
- ^ Robin Dennell and Wil Roebroeks (2003) "An Asian perspective on early human dispersal from Africa", Nature, Vol. 438, pp. 22-29. doi:10.1038/nature04259
- ^ Stringer, C. B. (2000) “Archaic Homo Sapiens”; in the Encyclopedia of Human Evolution and Prehistory, 2nd Ed (Eds: Delson, E. Tattersal, I. Van Couvering J. A. & Brooks, A. S.). Garland Publishing, NY. pp 159-168.
- ^ White, T. D. et al. (2003) "Pleistocene Homo sapiens from Middle Awash, Ethiopia", Nature. Vol. 423, pp. 742-747; doi:10.1038/nature01669
- ^ McDougall I. et al. (2005) "Stratigraphic placement and age of modern humans from Kibish, Ethiopia", Nature. Vol. 433, pp. 733-736; doi:10.1038/nature03258
- ^ Mellars, P. (2006) "Why did modern human populations disperse from Africa ca. 60,000 years ago? A new model" Proceedings of the National Academy of Sciences. Vol. 103 (25), pp. 9381-9386
- ^ Vanhaeren M. et al. (2005) "Middle Paleolithic Shell Beads in Israel and Algeria", Science. Vol. 312 (5781), pp. 1785-1788; doi:10.1126/science.1128139
- ^ Ian Tattersall (2008) "An Evolutionary Framework for the Acquisition of Symbolic Cognition by Homo sapiens", Comparative Cugnition and Behavior Reviews. Vol. 3, pp. 99-114; doi: 10.3819/ccbr.2008.30006
- ^ 1 2 3 Jorde, B. L. and Wooding S. P. (2004) "Genetic variation, classification and 'race'", Nature Genetics. Vol. 36, pp. S28 - S33; doi:10.1038/ng143
- ^ Long, J. C. and Kittles, R. A. (2003) "Human Genetic Diversity and the Nonexistence of Biological Races", Human Biology. Vol. 75, pp. 449-471
- ^ Hey, J. (2010) "The Divergence of Chimpanzee Species and Subspecies as Revealed in Multipopulation Isolation-with-Migration Analyses", Molecular Biology and Evolution. Vol. 27, pp. 921-933; doi: 10.1093/molbev/msp298
- ^ Gonder, M. K. et al. (2011) "Evidence from Cameroon reveals differences in the genetic structure and histories of chimpanzee populations", Proceedings of the National Academy of Sciences. DOI: 10.1073/pnas.1015422108
- ^ Hawks, J. et al. (2007) "Recent acceleration of human adaptive evolution", Proceedings of the National Academy of Sciences. Vol. 104 (52), pp.20753-20758 DOI: 10.1073/pnas.0707650104
- ^ Ingram, C. J. et al. (2009) "Lactose digestion and the evolutionary genetics of lactase persistence", Human Genetics. Vol. 124 (6), pp. 579-591. DOI: 10.1007/s00439-008-0593-6
- ^ Tishkof, A. S. (2006) "Convergent adaptation of human lactase persistence in Africa and Europe'" Nature Genetics. Vol. 39, pp. 31 - 40; doi:10.1038/ng1946
- ^ Mathieson I. et al. (2015) "Genome-wide patterns of selection in 230 ancient Europeans" Nature. doi:10.1038/nature16152
- ^ "Is Evolution a Theory or a Fact?" Evolution Resources From the National Academies
- ^ The Language of God, Prometheus Books, 2010, pp. 133-134
- ^ Kitzmiller v. Dover Area School District, Day 1 (September 26), AM Session, Part 2
- ^ The edge of Evolution, Free Press, 2007, pp. 71-72
- ^ אסף שטול-טראורינג, אדם המעוניין ללמוד על אודות הטבע, ילך אל הטבע, באתר הארץ, 19 במאי 2010
- ^ קובי נחשוני, הרשקוביץ: ייתכן מאוד שמוצא האדם מהקוף, באתר ynet, 4 במרץ 2010
- ^ "Public Acceptance of Evolution", Science
- ^ More Jews than Arabs believe in evolution, Pew Research Center, March 3, 2016
- ^ About half of Israeli Jews believe in evolution. Pew Research Center, March 3, 2016
- ^ שחר חי, אבולוציה בלי קוף: "נושא רגיש מאוד בישראל", באתר ynet, 1 ביוני 2014
- ^ Stephen J. Gould (1989) “Wonderful Life”; W. W. Norton & Company, NY. pp 23-38.
- ^ Donald R. Prothero, Carl Dennis Buell (2007) “Systematics and Evolution”; in Evolution: what the fossils say and why it matters. Columbia University Press, NY. pp 121-141.
